HS julkaisi puolitoista vuotta sitten Jani Kaaron artikkelin Tutkijat: Kovat rasvat aiheuttavat lihavuuteen liittyvän tulehduksen, joka käsitteli metaboliseen oireyhtymään liittyvää tulehdustilaa.[1]

Kaaro esitti useiden todisteiden viittaavan siihen että “tyydyttynyt rasva on se todellinen konna tulehdusreaktion takana” ja että syy on että se aktivoi elimistössä immuunijärjestelmään keskeisesti kuuluvia TLR-reseptoreita samalla tavalla kuin bakteerit, jolloin käynnistyy elimistöä rappeuttava tulehdustila. Tämä tulehdustila sitten johtaa insuliiniresistenssiin, korkeaan verenpaineeseen, keskivartalolihavuuteen ja lukuisiin muihin ongelmiin.
Tässä ollaan mielestäni jo vahvasti nykysairauksien syiden jäljillä. Kaaro on täysin oikeassa siinä että varsinkin TLR-reseptorien välityksellä syntyvä matala-asteinen tulehdustila on monien tutkijoiden mielestä keskeinen tekijä lukuisten kroonisten sairauksien taustalla.
Tässä olikin hyvä jutun alku, joten pohditaanpa ravinnon ja tulehduksen yhteyttä hieman perusteellisemmin!
2. Huono ruokavalio voi aiheuttaa haitallista endotoksemiaa
Aika ajoin rasva, hedelmäsokeri tai alkoholi saavat tietyissä piireissä huonon maineen koska nämä ruoka-aineet ovat etenkin erinäisissä eläintutkimuksissa aiheuttaneet metaboliseen oireyhtymään liittyviä ongelmia kuten tulehdusta, insuliiniresistenssiä ja dyslipidemiaa.
Näitä mainittuja ravintoaineita yhdistää se, että huonoon ruokavalioon yhdistettynä kyseiset ravintoaineet voivat aiheuttaa ns. metabolista endotoksemiaa, joka tarkoittaa yksinkertaisesti lisääntynyttä suolistobakteerien soluseinien lipopolysakkaridien (LPS) pääsyä verenkiertoon.
Endotoksemian määrä riippuu monesta tekijästä: Ruoka-aineet voivat vaikuttaa esimerkiksi suoliston bakteerikantaan, bakteerien määrään, suoliston seinämän läpäisevyyteen tai suoraan LPS:ien imeytymiseen. Myös erilaiset suolistossa tai veressä olevat tekijät kuten sappihapot, intestinal alkaline phosphatase (IAP) tai lipoproteiinit voivat vaikuttaa esimerkiksi LPS:n detoksifikaatioon.[2]
Miten elimistö reagoi endotoksemiaan? Tulehduksella.
Suuri osa näiden ravintoaineiden (rasva, sokeri, alkoholi) haitoista johtuu tulehduksesta, jonka aiheuttaa endotoksemia.
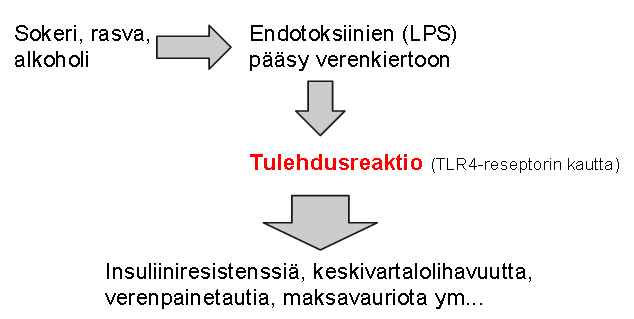 |
| 1) Huono ruokavalio voi aiheuttaa endotoksiinien pääsyä verenkiertoon. 2) Elimistö reagoi näihin endotoksiineihin tulehdusreaktiolla. 3) Tulehdus aiheuttaa metaboliseen oireyhtymään liittyvän ongelmien kirjon. |
Aiheesta on hyvin paljon näyttöä eläimillä, ja ihmisilläkin on havaittu lukuisia yhteyksiä endotoksemian ja metabolisen oireyhtymän välillä:[3-6]
- Hiirillä endotoksemian aiheuttaminen aiheuttaa lihomista, insuliiniresistenssiä ja kolesteroliongelmia
- Ihmisillä endotoksemian aiheuttaminen aiheuttaa rasvakudoksen tulehdusta ja insuliiniresistenssiä
Viime aikoina erinäisissä uutisissa hedelmäsokeria on pidetty osasyyllisenä maksaongelmiin kuten rasvamaksaan. Tämä on sinänsä loogista, sillä hedelmäsokeri voi ainakin tietyissä tilanteissa aiheuttaa ongelmia endotoksemian kautta.
Erään tutkimuksen mukaan rasvamaksapotilailla on veressä endotoksiinia lähes 30-kertaisesti verrattuna terveisiin ihmisiin. Rotilla jo pelkkä endotoksiinimäärän kaksinkertaistuminen riittää aiheuttamaan merkittäviä terveysongelmia.[6-8]
3. Tulehdusprosessi synnyttää ongelmat
Endotoksemia aiheuttaa elimistössä tulehdusprosessin, jota on eläinkokeissa kokeiltu estää tehokkaasti esimerkiksi lääkkein, geenimuunnoksilla tai tulehdussolujen poistolla.
Kun näin tehdään, muuttuvat sokeri, rasva ja alkoholi koe-eläimille hyvin harmittomiksi - tällöin esimerkiksi maksavauriota, lihavuutta ja insuliiniresistenssiä ei pääse yleensä syntymään suurillakaan annoksilla.[6,9-15]
Täten voidaan todeta että ongelmien todellinen aiheuttaja ei ole alkoholi, rasva, sokeri eikä vielä endotoksemiakaan vaan vasta tulehdusprosessin käynnistyminen synnyttää merkittävimmät ongelmat.
4. Lihavuus tukee tulehdusprosessia
Sen lisäksi, että elimistö reagoi suolistobakteereista johtuvaan endotoksemiaan tulehduksella, niin myös oman rasvakudoksen koko saattaa vaikuttaa kuvioon.
Erityisesti viskeraalisen rasvakudoksen kasvaessa tapahtuu kaksi asiaa:
- Rasvakudos erittää tulehdusproteiineja
- Rasvakudoksesta vapautuu rasvahappoja verenkiertoon (liiat rasvahapot voivat häiritä aineenvaihduntaa)
Nämä molemmat asiat voivat edistää kroonisten sairauksien kehittymistä.
 |
| Lihavuustutkija Stephan Guyenetin luennosta What Causes Leptin Resistance? (2014) |
5. Glysiini-aminohappo voi estää tulehduksen ja parantaa metabolisen oireyhtymän
Eläinkokeissa aiheeseen liittyy kiinnostavaa näyttöä eräästä yleisestä ravintoaineesta nimeltä glysiini (aminohappo). Glysiinin runsaampi syöminen hillitsee elimistössä tulehdusprosessia, joten se vähentää tulehduksen aiheuttamia haittoja.
Useissa tutkimuksissa on osoitettu että suuri määrä sokeria, alkoholia tai rasvaa voi aiheuttaa koe-eläimille rasvamaksaa, insuliiniresistenssiä ja hapetusstressiä. Glysiinin tai sitä runsaasti sisältävän proteiinin (kampasimpukka) lisääminen ruokavalioon estää nämä haitat lähes täydellisesti - suojaamalla suolistoa ja hillitsemällä tulehdusta.[16-20]
Annetaan vielä esimerkki eräästä tutkimuksesta:
Vuonna 2004 julkaistussa meksikolaistutkimuksessa rottien juomaveteen lisättiin 30% sokeria, jolloin rottien ainoa veden lähde sisälsi sokeria lähes kolminkertaisesti verrattuna tyypilliseen tuoremehuun tai virvoitusjuomaan. Tämän vuoksi rotat saivat vedenjuonnin ohessa niin valtavan määrän sokeria että varsinaiselle proteiinia, vitamiineja sekä mineraaleja sisältävälle ruoalle ei jäänyt juurikaan tilaa ja sen saanti vähenikin yli 50 prosentin verran.
Tuloksena syntynyt ravinneköyhä, äärimmäisen runsassokerinen ruokavalio aiheutti rotille huomattavaa verenpaineen nousua, keskivartalolihavuutta, veriarvojen huononemista sekä muita ongelmia.
Kuitenkin, jos juomaveteen lisättiin sokerin lisäksi myös 1% glysiiniä, lähes kaikki ravinneköyhän ja sokeripitoisen ruokavalion haitat kumoutuivat neljässä viikossa - vaikka rotat olivat noudattaneet haitallista ruokavaliota tätä ennen kaksikymmentä viikkoa eli viisinkertaisen ajan.[18]
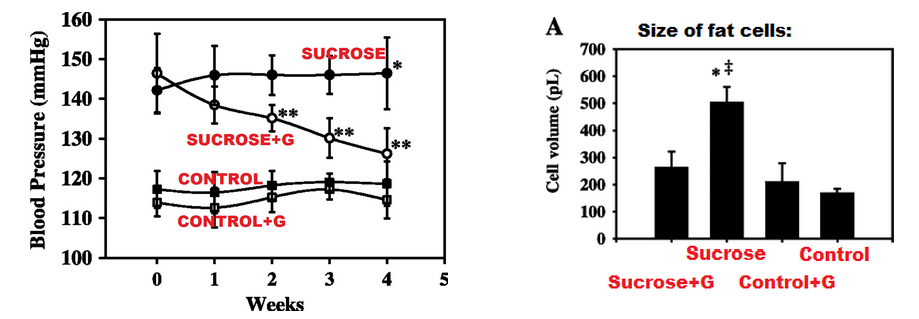 |
| Rottien verenpaineen lasku glysiinilisän myötä sekä rasvasolujen pienempi koko glysiiniryhmässä. (El Hafidi et al. 2004) |
Esimerkkinä mainitun tutkimuksen tulosta voisi olla vaikea uskoa todelliseksi, jollei sitä olisi toistettu muutamaankin otteeseen.
Myöhemmät eläintutkimukset ovat lähinnä osoittaneet, että glysiinin hyödyt eivät tosiaankaan rajoitu pelkästään verenpaineen alenemiseen ja rasvamassan vähenemiseen vaan ovat paljon kattavampia. Valitettavasti ihmistutkimuksissa tulokset ovat olleet pääosin maltillisempia.
6. Tulehdus sotkee energia-aineenvaihduntaa, mutta AMPK voi vielä pelastaa
Endotoksemian aiheuttamalla tulehduksella on elimistön soluihin erilaisia vaikutusmekanismeja, joiden kautta haittavaikutukset ilmenevät:
- Keramidien synteesi
- → NLRP3 → IL-1beta → Insuliiniresistenssi → Puutteita solujen energia-aineenvaihdunnassa
- Typpioksidi (iNOS)
- → Puutteita solujen energia-aineenvaihdunnassa
- Adiponektiinin aleneminen
- → Vähäisempi Sirt1/AMPK -aktivaatio → Puutteita solujen energia-aineenvaihdunnassa
- Vapaiden rasvahappojen tasojen nousu
- → DAG → PKC-theta → Insuliiniresistenssi → Puutteita solujen energia-aineenvaihdunnassa
Yllä olevat kohdat ovat vain viitteellisiä, mutta sanoma on silti se, että tulehdukseen liittyy useita mekanismeja jotka muuttavat solujen aineenvaihduntaa ja johtavat kroonisiin sairauksiin.
Näiltä haitoilta voi kuitenkin suojautua. Eläintutkimuksissa esimerkiksi erinäiset paastot ja aikarajoitetut syömismallit ovat suojanneet roskaruoan haitoilta. Ruokailun rajoittaminen 9 tai 12 tuntiin päivässä estää koe-eläimillä lukuisia huonon ruokavalion haittoja, vaikka kalorien kokonaissaanti ei muuttuisikaan.
Paastojen, joidenkin erikoisruokavalioiden (kuten ketogeenisen) sekä liikunnan hyötyjä on monesti perusteltu sillä, että ne aktivoivat solun energia-aineenvaihdunnan kannalta keskeistä AMPK-entsyymiä.
Erilaiset AMPK:ta aktivoivat aineet kuten AICAR, metformiini, koentsyymi Q10, aspiriini tai butyraatti, ovat koe-eläintutkimuksissa estäneet huomattavasti erilaisia roskaruokaan tai diabetekseen liittyviä haittoja. Tieteen uusin löytö lienee AICARin synteesiä lisäävä mitokondriopeptidi MOTS-c, joka estää koe-eläimillä lihavuutta ja muita roskaruoan aiheuttamia haittoja.
Täten tulehduksen estämisen lisäksi myös solujen energia-aineenvaihduntaan vaikuttaminen esimerkiksi AMPK:ta aktivoivilla menetelmillä tai ravintolisillä saattaisi suojata erinäisiltä kroonisilta sairauksilta. Jälleen kerran näyttäisi kuitenkin siltä, että vaikutukset ilmenevät koe-eläimillä hieman suurempina kuin ihmisillä.
7. Päätelmät
Kova rasva, sokeri tai alkoholi eivät usein aiheuta terveysongelmia suoraan, vaan ongelmat johtuvat ensisijaisesti elimistön omasta tulehdusprosessista, jota roskaruoka voi edistää lisäämällä endotoksiinien määrää veressä. Tulehdusprosessi taas voi sotkea solujen energia-aineenvaihduntaa.
Erilaiset ravintoaineet kuten glysiini, tauriini, antioksidantit, kuitu, maitohappobakteerit, histidiini*, karnosiini*, lyhytketjuiset rasvahapot, sinkki ja K-vitamiinit saattavat suojata endotoksemialta ja tulehdukselta vaikuttamalla esimerkiksi suolistoon tai tulehdussoluihin. Paastot, aikarajoitteinen syöminen, ketogeeninen ruokavalio, liikunta, ubikinoni, voihappo ja metformiini saattavat taas parantaa solujen energia-aineenvaihduntaa muun muassa AMPK-aktivaation kautta. Näyttöä on tosin eniten eläinkokeista, joita ei voi suoraan soveltaa ihmisiin.[21-47]
Antioksidanttien ja kuitujen endotoksemialta suojaava vaikutus voisi hypoteettisesti selittää, miksi Ruotsin maajusseilla runsas maitorasvan kulutus yhdistettynä runsaaseen kasvisten kulutukseen korreloi selvästi alentuneen sydäntautiriskin kanssa, vaikka maitorasva onkin pahamaineista "kovaa rasvaa".[48]
Lopuksi täytynee todeta kaiken näytön perusteella seuraavasti,
Jos keskittyisimme paremmin länsimaisten sairauksien mekanismeihin, voisimme estää sairaudet paremmin. Tällä hetkellä ravintokeskustelu junnaa usein pinnallisella tasolla, ja rasvan tai sokerin haittoja perustellaan muun muassa kolesteroliarvoilla tai verensokeritasojen heilahteluilla. Samalla tulehdusprosessin ja solusignaloinnin suuri merkitys kroonisten sairauksien taustalla jää helposti unohduksiin.
Lisämerkinnät
* Histidiini-aminohaposta ja karnosiinista vielä sellainen maininta, että eläinkoetulokset ovat näidenkin osalta vähintäänkin lupaavia.
Glysiinitutkimuksissa hyvin tehokas suoja sokerin tai alkoholin haittoja vastaan on saatu lisäämällä koe-eläinten juomaveteen 1% glysiiniä eli 10 grammaa litraa kohden. Näiden kahden ravinteen kohdalla jo 0.1% eli 1 gramma vesilitraa kohden näyttää taas tuottavan kohtalaisen suuria hyötyjä.
Eräässä kohtalaisen suuressa ihmistutkimuksessa ylipainosta kärsivät naiset söivät päivittäin 4 gramman annoksen histidiiniä (melko suuri annos), mikä johti 12 viikon aikana merkittävään tulehdusmarkkerien laskuun, insuliiniherkkyyden paranemiseen sekä lähes kolmen kilon vähennykseen rasvamassassa. Tarkasta vaikutusmekanismista minulla ei ole tietoa, mutta vaikutus saattaa liittyä mm. karnosiinipitoisuuden nousuun ja esimerkiksi sen lipidiperoksideilta suojaavaan vaikutukseen.[33,49]
Mainittakoon myös että histidiini ja karnosiini nostavat toistensa veritasoja, ja että karnosiini on dipeptidi joka koostuu histidiinistä sekä beeta-alaniinista. Karnosiinia pidetään tietyissä piireissä hyvin terveellisenä ravinteena, ja esimerkiksi Tohtori Tolonen on mainostanut sitä Suomessa hyvin paljon. Eläinkokeiden perusteella histidiini ja karnosiini ovat suunnilleen yhtä tehokkaita, mutta histidiini on selkeästi halvempi tuote, erityisesti jauheena (joskin vaikeammin saatavilla - esimerkiks iHerb ei näytä myyvän jauhemuotoa lainkaan).
Lisämateriaali, I: Videoita ja esitelmiä
Lihavuustutkija Stephan Guyenetin tulehdusta ja lihavuutta käsittelevä luento Ancestral Health Symposium 2014 -tapahtumasta. Todella laadukasta katsottavaa, ja näkemykset hyvin samanlaiset kuin minulla.
Seuraavan kuvan Guyenetin luennosta jätän tähän kertausta varten:

Guyenet näyttää, että tulehdus sekä korkea leptiini voivat erityisesti SOCS3:n kautta häiritä leptiinin STAT3-välitteistä signalointia solussa.
Samoin runsas määrä rasvametaboliitteja (diasyyliglyserolit ja Acyl-CoA) solun sisällä voivat PKC-thetan kautta häiritä leptiinin PI3K-välitteistä signalointia (palmitiinihappo lisää rasvametaboliitteja enemmän kuin oleiinihappo). Lisäksi tulehdus häiritsee leptiinin STAT3-välitteistä signalointia myös PTP1B:n kautta, ja liikunta taas voi parantaa leptiiniherkkyyttä vähentämällä PTP1B-aktiivisuutta IL-6 -sytokiinin kautta.
Upgraded Life Festival -tapahtumassa esittämäni glysiini-esitelmän luentodiat, jossa esitellään glysiinin vaikutuksia lyhyesti mutta laajasti.
Lisämateriaali, II: Kova rasva ja suora TLR4-aktivaatio
Olen käsittänyt että kovan rasvan aiheuttama tulehdus on ensisijaisesti epäsuoraa eli endotoksemian kautta aiheutuvaa.
On kuitenkin olemassa runsaasti julkaisuja, joissa näytetään että kova rasva voi mm. koeputkessa itsessäänkin aktivoida kyseistä reseptoria.[53-57] Ilmeisesti Kaaron artikkelissa tarkoitetaan juuri tätä mekanismia. Ylipainoisilla ihmisillä on enemmän rasvakudosta, jonka vapauttama kova rasva voisi itsessäänkin aiheuttaa tulehdusta.
Mikäli tämä mekanismi on myös keskeinen kovan rasvan haitoille, niin se saattaisi myös osittain selittää glysiinin tai histidiinin suurta tehoa joissakin tutkimuksissa. Eläinkokeissa glysiini hillitsee rasvakudoksen tulehdusta, ja jos rasvakudoksen tulehdus on merkittävä ongelmien lähde, niin se varmasti selittää vaikutuksia jonkun verran.[58-60]
Vaikka lähinnä kova rasva näyttäisikin aktivoivan TLR4-reseptoria suorasti, en tiedä tekeekö se itsessään kovasta rasvasta jotenkin "huonomman" rasvan. Koeputkitason tutkimuksissa on myös pehmeistä rasvoista huomattu haitallisia puolia kuten mm. aivojen turpoaminen. On myös kymmenittäin eläintutkimuksia, joissa kovan rasvan syöttäminen suojaa pehmeän rasvan aiheuttamalta maksavauriolta.[61-66]
Näiden havaintojen pohjalta olen oikeastaan miettinyt, että kova rasva (voi, kookosrasva) voisi olla lähtökohtaisesti jopa terveellisin rasvan lähde sillä siihen ei liity monia pehmeisiin rasvoihin liittyviä biokemiallisia ongelmia, mutta toisaalta kontekstin on oltava sellainen jossa terveellinen ruokavalio (ja esim glysiini) minimoivat TLR4-välitteisen tulehdusreaktion haitat.
Lisämateriaali, III: Lisäpohdintaa sairauksien mekanismeista
Metabolisen oireyhtymän syymekanismeja ja hoitoa pohdittaessa voi TLR4-aktivaatioon liittyvien tulehdusmekanismien lisäksi kiinnittää huomion voi esimerkiksi kiinnittää myös kortisoliaineenvaihduntaan (HSD-11β -entsyymit), mitokondrioiden toimintaan, solujen antioksidantteihin, lipidiperoksidaatioon, prostaglandiiniaineenvaihduntaan (COX-2, PGE2...), vapaiden rasvahappojen fysiologisiin vaikutuksiin (NEFA/FFA) sekä kilpirauhashormonien vaikutuksiin. Itse näen mm. Robert Naviauxin ja Ray Peatin vaihtoehtoiset näkökulmat hyvin kiinnostavina kokonaiskuvan kannalta.
Suoliston terveys on olennaisessa roolissa kun pohditaan endotoksemian ehkäisyä. Alentunut aineenvaihdunta (esim fibromyalgiassa tai kilpirauhasen vajaatoiminnassa) voinee suolen toimintaa hidastamalla lisätä bakteerien kasvua ohutsuolessa, mikä vaikuttaisi haitallisesti ohutsuoleen ja täten pahentaisi myös endotoksemiaa. En ole vielä perehtynyt aiheeseen kunnolla, mutta tähän voisin kesän aikana yrittää paneutua.[50-52]
Lähteitä
[1] HS 17.7.2012, Jani Kaaro: Tutkijat: Kovat rasvat aiheuttavat lihavuuteen liittyvän tulehduksen
[2] J Mol Endocrinol. 2013 Sep 11;51(2):R51-64. Metabolic endotoxemia: a molecular link between obesity and cardiovascular risk. Neves AL, Coelho J, Couto L, Leite-Moreira A, Roncon-Albuquerque R Jr.
[3] Diabetes. 2007 Jul;56(7):1761-72. Metabolic endotoxemia initiates obesity and insulin resistance. Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, Neyrinck AM, Fava F, Tuohy KM, Chabo C, Waget A, Delmée E, Cousin B, Sulpice T, Chamontin B, Ferrières J, Tanti JF, Gibson GR, Casteilla L, Delzenne NM, Alessi MC, Burcelin R.
[4] Diabetes. 2010 Jan;59(1):172-81. Experimental endotoxemia induces adipose inflammation and insulin resistance in humans. Mehta NN, McGillicuddy FC, Anderson PD, Hinkle CC, Shah R, Pruscino L, Tabita-Martinez J, Sellers KF, Rickels MR, Reilly MP.
[5] J Transl Med. 2012 Jun 18;10:124. A human model of inflammatory cardio-metabolic dysfunction; a double blind placebo-controlled crossover trial. Mehta NN, Heffron SP, Patel PN, Ferguson J, Shah RD, Hinkle CC, Krishnamoorthy P, Shah R, Tabita-Martinez J, Terembula K, Master SR, Rickels MR, Reilly MP.
[6] Am J Physiol Endocrinol Metab. 2007 Mar;292(3):E740-7. Lipopolysaccharide activates an innate immune system response in human adipose tissue in obesity and type 2 diabetes. Creely SJ, McTernan PG, Kusminski CM, Fisher fM, Da Silva NF, Khanolkar M, Evans M, Harte AL, Kumar S.
[7a] HS 24.6.2013, Jani Kaaro: Tutkimus: Hedelmäsokeri aiheuttaa rasvamaksaa ilman painonnousuakin
[7b] Am J Clin Nutr. 2013 Aug;98(2):349-57. doi: 10.3945/ajcn.112.057331. Dietary fructose induces endotoxemia and hepatic injury in calorically controlled primates. Kavanagh K, Wylie AT, Tucker KL, Hamp TJ, Gharaibeh RZ, Fodor AA, Cullen JM.
[8] J Nutr. 2008 Aug;138(8):1452-5. Nonalcoholic fatty liver disease in humans is associated with increased plasma endotoxin and plasminogen activator inhibitor 1 concentrations and with fructose intake. Thuy S, Ladurner R, Volynets V, Wagner S, Strahl S, Königsrainer A, Maier KP, Bischoff SC, Bergheim I.
[9] Diabetes. 2010 Feb;59(2):347-57. doi: 10.2337/db09-0016. Epub 2009 Nov 23. Depletion of liver Kupffer cells prevents the development of diet-induced hepatic steatosis and insulin resistance. Huang W, Metlakunta A, Dedousis N, Zhang P, Sipula I, Dube JJ, Scott DK, O'Doherty RM.
[10] Methods Mol Biol. 2008;447:199-212. doi: 10.1007/978-1-59745-242-7_15. Isolation of Kupffer cells from rats fed chronic ethanol. McMullen MR, Pritchard MT, Nagy LE.
[11] Diabetes. 2007 Aug;56(8):1986-98. Epub 2007 May 22. Loss-of-function mutation in Toll-like receptor 4 prevents diet-induced obesity and insulin resistance. Tsukumo DM, Carvalho-Filho MA, Carvalheira JB, Prada PO, Hirabara SM, Schenka AA, Araújo EP, Vassallo J, Curi R, Velloso LA, Saad MJ.
[12] Hepatology. 1994 Aug;20(2):453-60. Inactivation of Kupffer cells prevents early alcohol-induced liver injury. Adachi Y, Bradford BU, Gao W, Bojes HK, Thurman RG.
[13] J Hepatol. 2008 Jun;48(6):983-92. doi: 10.1016/j.jhep.2008.01.035. Epub 2008 Mar 14. Antibiotics protect against fructose-induced hepatic lipid accumulation in mice: role of endotoxin. Bergheim I, Weber S, Vos M, Krämer S, Volynets V, Kaserouni S, McClain CJ, Bischoff SC. "Total caloric intake and weight gain were highest in mice exposed to glucose."
[14] Gastroenterology. 1995 Jan;108(1):218-24. Antibiotics prevent liver injury in rats following long-term exposure to ethanol. Adachi Y1, Moore LE, Bradford BU, Gao W, Thurman RG.
[15] Diabetes. 2008 Jun;57(6):1470-81. doi: 10.2337/db07-1403. Epub 2008 Feb 27. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Cani PD, Bibiloni R, Knauf C, Waget A, Neyrinck AM, Delzenne NM, Burcelin R. "Furthermore, the absence of CD14 in ob/ob CD14(-)(/)(-) mutant mice mimicked the metabolic and inflammatory effects of antibiotics."
[16] Amino Acids. 2014 Mar 23. [Epub ahead of print] Scallop protein with endogenous high taurine and glycine content prevents high-fat, high-sucrose-induced obesity and improves plasma lipid profile in male C57BL/6J mice. Tastesen HS, Keenan AH, Madsen L, Kristiansen K, Liaset B.
[17] Clin Sci (Lond). 2014 Jan 1;126(1):19-29. doi: 10.1042/CS20130164. Glycine restores glutathione and protects against oxidative stress in vascular tissue from sucrose-fed rats. Ruiz-Ramírez A, Ortiz-Balderas E, Cardozo-Saldaña G, Diaz-Diaz E, El-Hafidi M.
[18] Am J Physiol Regul Integr Comp Physiol. 2004 Dec;287(6):R1387-93. Epub 2004 Aug 26. Glycine intake decreases plasma free fatty acids, adipose cell size, and blood pressure in sucrose-fed rats. El Hafidi M, Pérez I, Zamora J, Soto V, Carvajal-Sandoval G, Baños G.
[19] Can J Physiol Pharmacol. 2011 Dec;89(12):899-910. doi: 10.1139/y11-086. Epub 2011 Nov 24. Effect of glycine on the cyclooxygenase pathway of the kidney arachidonic acid metabolism in a rat model of metabolic syndrome. Pérez-Torres I1, Ibarra B, Soria-Castro E, Torrico-Lavayen R, Pavón N, Diaz-Diaz E, Flores PL, Infante O, Baños G.
[20] Cell Biochem Funct. 2004 Mar-Apr;22(2):123-8. Protective effect of glycine supplementation on the levels of lipid peroxidation and antioxidant enzymes in the erythrocyte of rats with alcohol-induced liver injury. Senthilkumar R, Sengottuvelan M, Nalini N
[21] Am J Clin Nutr. 2010 Apr;91(4):940-9. Orange juice neutralizes the proinflammatory effect of a high-fat, high-carbohydrate meal and prevents endotoxin increase and Toll-like receptor expression. Ghanim H, Sia CL, Upadhyay M, Korzeniewski K, Viswanathan P, Abuaysheh S, Mohanty P, Dandona P.
[22] J Clin Endocrinol Metab. 2011 May;96(5):1409-14. A resveratrol and polyphenol preparation suppresses oxidative and inflammatory stress response to a high-fat, high-carbohydrate meal. Ghanim H, Sia CL, Korzeniewski K, Lohano T, Abuaysheh S, Marumganti A, Chaudhuri A, Dandona P.
[23] Endocrine Abstracts (2012) The intake of fiber suppresses the high fat high carbohydrate meal induced endotoxemia, oxidative stress and inflammation H. Ghanim, M. Batra, S. Dhindsa, K. Green, S. Abuaysheh, A. Chaudhuri & P. Dandona
[24] Br J Nutr. 2009 Apr;101(7):961-6. Probiotic yogurt in the elderly with intestinal bacterial overgrowth: endotoxaemia and innate immune functions. Schiffrin EJ1, Parlesak A, Bode C, Bode JC, van't Hof MA, Grathwohl D, Guigoz Y.
[25] Nutrition. 2012 May;28(5):539-43. Probiotic yogurt improves antioxidant status in type 2 diabetic patients. Ejtahed HS1, Mohtadi-Nia J, Homayouni-Rad A, Niafar M, Asghari-Jafarabadi M, Mofid V. "Probiotic yogurt significantly decreased fasting blood glucose (P < 0.01) and hemoglobin A1c (P < 0.05) and increased erythrocyte superoxide dismutase and glutathione peroxidase activities and total antioxidant status (P < 0.05) compared with the control group [conventional yogurt]."
[26] Ann Nutr Metab. 2013;63(1-2):1-9. Effect of multispecies probiotic supplements on metabolic profiles, hs-CRP, and oxidative stress in patients with type 2 diabetes. Asemi Z1, Zare Z, Shakeri H, Sabihi SS, Esmaillzadeh A. "prevented a rise in FPG and resulted in a decrease in serum hs-CRP and an increase in plasma total GSH"
[27] Iran J Pharm Res. 2013 Fall;12(4):929-36. Probiotic yogurt Affects Pro- and Anti-inflammatory Factors in Patients with Inflammatory Bowel Disease. Shadnoush M, Shaker Hosseini R, Mehrabi Y, Delpisheh A, Alipoor E, Faghfoori Z, Mohammadpour N, Zaringhalam Moghadam J.
[28] Biochem Biophys Res Commun. 2013 Feb 8;431(2):258-63. Lactobacillus rhamnosus GG improves insulin sensitivity and reduces adiposity in high-fat diet-fed mice through enhancement of adiponectin production. Kim SW, Park KY, Kim B, Kim E, Hyun CK.
[29] PLoS One. 2014 Jan 27;9(1):e80169. Lactobacillus rhamnosus GG protects against non-alcoholic fatty liver disease in mice. Ritze Y1, Bárdos G1, Claus A1, Ehrmann V1, Bergheim I2, Schwiertz A3, Bischoff SC1.
[30] Am J Pathol. 2011 Dec;179(6):2866-75. Lactobacillus rhamnosus GG treatment potentiates intestinal hypoxia-inducible factor, promotes intestinal integrity and ameliorates alcohol-induced liver injury. Wang Y1, Kirpich I, Liu Y, Ma Z, Barve S, McClain CJ, Feng W.
[31] J Nutr Biochem. 2013 Sep;24(9):1609-15. Lactobacillus rhamnosus GG reduces hepatic TNFα production and inflammation in chronic alcohol-induced liver injury. Wang Y1, Liu Y, Kirpich I, Ma Z, Wang C, Zhang M, Suttles J, McClain C, Feng W.
[32] World J Gastroenterol. 2013 Jan 14;19(2):274-83. Dual probiotic strains suppress high fructose-induced metabolic syndrome. Park DY, Ahn YT, Huh CS, McGregor RA, Choi MS.
[33] Diabetologia. 2013 May;56(5):985-94. Histidine supplementation improves insulin resistance through suppressed inflammation in obese women with the metabolic syndrome: a randomised controlled trial. Feng RN, Niu YC, Sun XW, Li Q, Zhao C, Wang C, Guo FC, Sun CH, Li Y.
[34] Eur J Pharmacol. 2011 Feb 25;653(1-3):82-8. Histidine and carnosine alleviated hepatic steatosis in mice consumed high saturated fat diet. Mong MC1, Chao CY, Yin MC.
[35] Eur J Pharmacol. 2005 Apr 18;513(1-2):145-50. Histidine and carnosine delay diabetic deterioration in mice and protect human low density lipoprotein against oxidation and glycation. Lee YT, Hsu CC, Lin MH, Liu KS, Yin MC. "1 g/l histidine and carnosine treatments significantly reduced cholesterol level in heart and liver (P < 0.05). The administration of histidine or carnosine significantly enhanced catalase activity and decreased lipid oxidation levels in kidney and liver (P < 0.05); however, only 1 g/l histidine and carnosine treatments significantly increased glutathione peroxidase activity (P < 0.05). The increased interleukin (IL)-6 and tumor necrosis factor (TNF)-alpha in diabetic mice were significantly suppressed by the intake of histidine or carnosine (P < 0.05)."
[36] Food Chem Toxicol. 2008 May;46(5):1503-9. Beneficial effects of histidine and carnosine on ethanol-induced chronic liver injury. Liu WH, Liu TC, Yin MC.
[37] J Food Sci. 2009 Oct;74(8):H259-65. Protective effects from carnosine and histidine on acetaminophen-induced liver injury. Yan SL, Wu ST, Yin MC, Chen HT, Chen HC.
[38] Tohoku J Exp Med. 2000 Jun;191(2):85-99. Effect of carnosine on rats under experimental brain ischemia. Gallant S, Kukley M, Stvolinsky S, Bulygina E, Boldyrev A.
[39] Nutr Res Pract. 2011 Oct;5(5):421-8. Effects of α-lipoic acid and L-carnosine supplementation on antioxidant activities and lipid profiles in rats. Kim MY1, Kim EJ, Kim YN, Choi C, Lee BH.
[40] PLoS One. 2011 Mar 15;6(3):e17971. Effects of dietary supplementation of carnosine on mitochondrial dysfunction, amyloid pathology, and cognitive deficits in 3xTg-AD mice. Corona C, Frazzini V, Silvestri E, Lattanzio R, La Sorda R, Piantelli M, Canzoniero LM, Ciavardelli D, Rizzarelli E, Sensi SL.
[41] J Physiol Biochem. 2014 Jun;70(2):385-95. Effect of carnosine alone or combined with α-tocopherol on hepatic steatosis and oxidative stress in fructose-induced insulin-resistant rats. Giriş M, Doğru-Abbasoğlu S, Kumral A, Olgaç V, Koçak-Toker N, Uysal M.
[42] Diabetes. 2009 Jul;58(7):1509-17. doi: 10.2337/db08-1637. Epub 2009 Apr 14. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Gao Z, Yin J, Zhang J, Ward RE, Martin RJ, Lefevre M, Cefalu WT, Ye J. "On the high-fat diet, supplementation of butyrate prevented development of insulin resistance and obesity in C57BL/6 mice."
[43] Am J Pathol. 2004 May;164(5):1547-56. Abrogation of nuclear factor-kappaB activation is involved in zinc inhibition of lipopolysaccharide-induced tumor necrosis factor-alpha production and liver injury. Zhou Z, Wang L, Song Z, Saari JT, McClain CJ, Kang YJ.
[44] J Pharmacol Exp Ther. 2003 Jun;305(3):880-6. Epub 2003 Mar 6. Prevention of alterations in intestinal permeability is involved in zinc inhibition of acute ethanol-induced liver damage in mice. Lambert JC, Zhou Z, Wang L, Song Z, McClain CJ, Kang YJ.
[45] Proc Natl Acad Sci U S A. 2013 Apr 23;110(17):7003-8. Intestinal alkaline phosphatase prevents metabolic syndrome in mice. Kaliannan K, Hamarneh SR, Economopoulos KP, Nasrin Alam S, Moaven O, Patel P, Malo NS, Ray M, Abtahi SM, Muhammad N, Raychowdhury A, Teshager A, Mohamed MM, Moss AK, Ahmed R, Hakimian S, Narisawa S, Millán JL, Hohmann E, Warren HS, Bhan AK, Malo MS, Hodin RA.
[46] J Nutr Sci Vitaminol (Tokyo). 2007 Jun;53(3):219-24. Enhancement effects of vitamin K1 (phylloquinone) or vitamin K2 (menaquinone-4) on intestinal alkaline phosphatase activity in rats. Sogabe N1, Maruyama R, Hosori T, Goseki-Sone M.
[47] Bone. 2011 May 1;48(5):1036-42. Effects of long-term vitamin K(1) (phylloquinone) or vitamin K(2) (menaquinone-4) supplementation on body composition and serum parameters in rats. Sogabe N1, Maruyama R, Baba O, Hosoi T, Goseki-Sone M.
[48] Int J Environ Res Public Health. 2009 Oct;6(10):2626-38. Food choices and coronary heart disease: a population based cohort study of rural Swedish men with 12 years of follow-up. Holmberg S, Thelin A, Stiernström EL.
[49] J Neurochem. 2007 May;101(3):729-36. Epub 2007 Jan 24. Neuroprotective actions of a histidine analogue in models of ischemic stroke. Tang SC, Arumugam TV, Cutler RG, Jo DG, Magnus T, Chan SL, Mughal MR, Telljohann RS, Nassar M, Ouyang X, Calderan A, Ruzza P, Guiotto A, Mattson MP. "We recently synthesized and characterized histidine analogues related to the natural dipeptide carnosine, which selectively scavenge the toxic lipid peroxidation product 4-hydroxynonenal (HNE)[...]"
[50] J Clin Endocrinol Metab. 2007 Nov;92(11):4180-4. Epub 2007 Aug 14. Association between hypothyroidism and small intestinal bacterial overgrowth. Lauritano EC, Bilotta AL, Gabrielli M, Scarpellini E, Lupascu A, Laginestra A, Novi M, Sottili S, Serricchio M, Cammarota G, Gasbarrini G, Pontecorvi A, Gasbarrini A.
[51] Ann Rheum Dis. 2004 Apr;63(4):450-2. A link between irritable bowel syndrome and fibromyalgia may be related to findings on lactulose breath testing. Pimentel M, Wallace D, Hallegua D, Chow E, Kong Y, Park S, Lin HC.
[52] Am J Gastroenterol. 2002 Sep;97(9):2364-70. Small intestinal bacterial overgrowth in human cirrhosis is associated with systemic endotoxemia. Bauer TM, Schwacha H, Steinbrückner B, Brinkmann FE, Ditzen AK, Aponte JJ, Pelz K, Berger D, Kist M, Blum HE.
[53] Br J Nutr. 2012 Jan;107(2):229-41. doi: 10.1017/S0007114511002868. Epub 2011 Jun 29. Saturated fatty acids activate microglia via Toll-like receptor 4/NF-κB signalling. Wang Z, Liu D, Wang F, Liu S, Zhao S, Ling EA, Hao A.
[54] J Biol Chem. 2001 May 18;276(20):16683-9. Epub 2001 Mar 2. Saturated fatty acids, but not unsaturated fatty acids, induce the expression of cyclooxygenase-2 mediated through Toll-like receptor 4. Lee JY, Sohn KH, Rhee SH, Hwang D.
[55] Arterioscler Thromb Vasc Biol. 2007 Jan;27(1):84-91. Epub 2006 Nov 2. Role of the Toll-like receptor 4/NF-kappaB pathway in saturated fatty acid-induced inflammatory changes in the interaction between adipocytes and macrophages. Suganami T, Tanimoto-Koyama K, Nishida J, Itoh M, Yuan X, Mizuarai S, Kotani H, Yamaoka S, Miyake K, Aoe S, Kamei Y, Ogawa Y. "These findings suggest that saturated FAs, which are released in large quantities from hypertrophied adipocytes via the macrophage-induced adipocyte lipolysis, serve as a naturally occurring ligand for TLR4, thereby inducing the inflammatory changes in both adipocytes and macrophages through NF-kappaB activation."
[56] J Neurochem. 2012 Mar;120(6):1060-71. doi: 10.1111/j.1471-4159.2012.07660.x. Epub 2012 Feb 6. Saturated long-chain fatty acids activate inflammatory signaling in astrocytes. Gupta S, Knight AG, Gupta S, Keller JN, Bruce-Keller AJ. "Data show that the saturated fatty acid palmitic acid, as well as lauric acid and stearic acid, trigger the release of TNFα and IL-6 from astrocytes. Unsaturated fatty acids were unable to induce cytokine release from cultured astrocytes. Furthermore, the effects of palmitic acid on cytokine release require Toll-like receptor 4 rather than CD36 or Toll-like receptor 2, and do not depend on palmitic acid metabolism to palmitoyl-CoA. [...] Finally, data show that the essential ω-3 fatty acid docosahexaenoic acid acts in a dose-dependent manner to prevent the actions of palmitic acid on inflammatory signaling in astrocytes. Collectively, these data demonstrate the ability of saturated fatty acids to induce astrocyte inflammation in vitro."
[57] J Neurosci. 2009 Jan 14;29(2):359-70. doi: 10.1523/JNEUROSCI.2760-08.2009. Saturated fatty acids produce an inflammatory response predominantly through the activation of TLR4 signaling in hypothalamus: implications for the pathogenesis of obesity. Milanski M, Degasperi G, Coope A, Morari J, Denis R, Cintra DE, Tsukumo DM, Anhe G, Amaral ME, Takahashi HK, Curi R, Oliveira HC, Carvalheira JB, Bordin S, Saad MJ, Velloso LA. "According to our results, long-chain saturated fatty acids activate predominantly toll-like receptor 4 signaling, which determines not only the induction of local cytokine expression but also promotes endoplasmic reticulum stress. Rats fed on a monounsaturated fat-rich diet do not develop hypothalamic leptin resistance, whereas toll-like receptor 4 loss-of-function mutation and immunopharmacological inhibition of toll-like receptor 4 protects mice from diet-induced obesity"
[58] Biomed Pharmacother. 2010 Oct;64(8):534-40. Glycine regulates inflammatory markers modifying the energetic balance through PPAR and UCP-2. Almanza-Perez JC, Alarcon-Aguilar FJ, Blancas-Flores G, Campos-Sepulveda AE, Roman-Ramos R, Garcia-Macedo R, Cruz M.
[59] Eur J Pharmacol. 2008 Jun 10;587(1-3):317-21. Glycine increases mRNA adiponectin and diminishes pro-inflammatory adipokines expression in 3T3-L1 cells. Garcia-Macedo R, Sanchez-Muñoz F, Almanza-Perez JC, Duran-Reyes G, Alarcon-Aguilar F, Cruz M.
[60] Eur J Pharmacol. 2008 Dec 3;599(1-3):152-8. Glycine regulates the production of pro-inflammatory cytokines in lean and monosodium glutamate-obese mice. Alarcon-Aguilar FJ, Almanza-Perez J, Blancas G, Angeles S, Garcia-Macedo R, Roman R, Cruz M.
[61] Science. 1978 Jul 28;201(4353):358-60. Brain edema: induction in cortical slices by polyunsaturated fatty acids. Chan PH, Fishman RA.
[62] Ann Neurol. 1983 Jun;13(6):625-32. Induction of brain edema following intracerebral injection of arachidonic acid. Chan PH, Fishman RA, Caronna J, Schmidley JW, Prioleau G, Lee J.
[63] Nutr Metab (Lond). 2007 Feb 21;4:4. Long term highly saturated fat diet does not induce NASH in Wistar rats. Romestaing C, Piquet MA, Bedu E, Rouleau V, Dautresme M, Hourmand-Ollivier I, Filippi C, Duchamp C, Sibille B.
[64] Alcohol. 2013 May;47(3):257-64. Ethanol and dietary unsaturated fat (corn oil/linoleic acid enriched) cause intestinal inflammation and impaired intestinal barrier defense in mice chronically fed alcohol. Kirpich IA, Feng W, Wang Y, Liu Y, Beier JI, Arteel GE, Falkner KC, Barve SS, McClain CJ.
[65] Hepatology. 2005 Sep;42(3):568-77. Role of adiponectin in the protective action of dietary saturated fat against alcoholic fatty liver in mice. You M, Considine RV, Leone TC, Kelly DP, Crabb DW.
[66] J Pharmacol Exp Ther. 2001 Nov;299(2):638-44. Dietary saturated fatty acids reverse inflammatory and fibrotic changes in rat liver despite continued ethanol administration. Nanji AA, Jokelainen K, Tipoe GL, Rahemtulla A, Dannenberg AJ.
Ylimääräisiä lähteitä (sekalaista aiheeseen liittyvää tutkimuskirjallisuutta)
[extra] Bol Med Hosp Infant Mex Vol. 67, March-April 2010 Obesity as an inflammatory process. Blancas-Flores G, Almanza-Pérez JC, López-Roa RI, Alarcón-Aguilar FJ, García-Macedo R, Cruz M.
[extra] EMBO Mol Med. 2011 Sep;3(9):559-72. doi: 10.1002/emmm.201100159. Epub 2011 Aug 3. Intestinal mucosal adherence and translocation of commensal bacteria at the early onset of type 2 diabetes: molecular mechanisms and probiotic treatment. Amar J, Chabo C, Waget A, Klopp P, Vachoux C, Bermúdez-Humarán LG, Smirnova N, Bergé M, Sulpice T, Lahtinen S, Ouwehand A, Langella P, Rautonen N, Sansonetti PJ, Burcelin R. "Here, we demonstrate that before the onset of diabetes, after only one week of a high-fat diet (HFD), live commensal intestinal bacteria are present in large numbers in the adipose tissue and the blood where they can induce inflammation. This translocation is prevented in mice lacking the microbial pattern recognition receptors Nod1 or CD14, but overtly increased in Myd88 knockout and ob/ob mouse. [...] The bacterial translocation process from intestine towards tissue can be reversed by six weeks of treatment with the probiotic strain Bifidobacterium animalis subsp. lactis 420, which improves the animals' overall inflammatory and metabolic status."
[extra] PLoS One. 2014 May 14;9(5):e96864. doi: 10.1371/journal.pone.0096864. eCollection 2014. Acute binge drinking increases serum endotoxin and bacterial DNA levels in healthy individuals. Bala S, Marcos M, Gattu A, Catalano D, Szabo G.
[extra] Alcohol Clin Exp Res. 2005 Nov;29(11 Suppl):166S-71S. Activation of the innate immune system and alcoholic liver disease: effects of ethanol per se or enhanced intestinal translocation of bacterial toxins induced by ethanol? Bode C, Bode JC.
[extra] J Hepatol. 1991 Mar;12(2):162-9. Plasma endotoxin concentrations in patients with alcoholic and non-alcoholic liver disease: reevaluation with an improved chromogenic assay. Fukui H, Brauner B, Bode JC, Bode C. "The results indicate that, irrespective of the stage of liver disease, alcohol abuse favours the development of endotoxemia."
[extra] J Hepatol. 2000 May;32(5):742-7. Increased intestinal permeability to macromolecules and endotoxemia in patients with chronic alcohol abuse in different stages of alcohol-induced liver disease. Parlesak A, Schäfer C, Schütz T, Bode JC, Bode C. "Polyethylene glycol Mr 10,000 was detected significantly less frequently in urine from healthy controls (0/30) than in urine of patients with alcoholic liver disease (20/54, [...] Endotoxin concentrations in the plasma of alcoholics were increased more than 5-fold compared to healthy controls [...] The results of this study indicate that alcohol abuse impairs the function of the intestinal barrier, which might enhance the translocation of bacterial toxins, thereby contributing to inflammatory processes in alcoholic liver disease."
[extra] Am J Clin Nutr. 2008 May;87(5):1219-23. Energy intake is associated with endotoxemia in apparently healthy men. Amar J, Burcelin R, Ruidavets JB, Cani PD, Fauvel J, Alessi MC, Chamontin B, Ferriéres J. "In a multivariate analysis, endotoxemia was independently associated with energy intake."
[extra] Diabetes Care. 2011 Feb;34(2):392-7. doi: 10.2337/dc10-1676. Endotoxemia is associated with an increased risk of incident diabetes. Pussinen PJ, Havulinna AS, Lehto M, Sundvall J, Salomaa V.
[extra] Nutr Metab (Lond). 2013 Jan 10;10(1):6. Dietary oil composition differentially modulates intestinal endotoxin transport and postprandial endotoxemia. Mani V, Hollis JH, Gabler NK.
[extra] Exp Physiol. 2014 Jun 27. [Epub ahead of print] Fructose supplementation worsens the deleterious effects of short term high fat feeding on hepatic steatosis and lipid metabolism in adult rats. Crescenzo R, Bianco F, Coppola P, Mazzoli A, Tussellino M, Carotenuto R, Liverini G, Iossa S.
[extra] Alcohol Clin Exp Res. 2012 May;36(5):835-46. doi: 10.1111/j.1530-0277.2011.01673.x. Epub 2011 Dec 7. The type of dietary fat modulates intestinal tight junction integrity, gut permeability, and hepatic toll-like receptor expression in a mouse model of alcoholic liver disease. Kirpich IA, Feng W, Wang Y, Liu Y, Barker DF, Barve SS, McClain CJ. "Compared with control animals, hepatic TLR (TLR 1, 2, 3, 4, 7, 8, 9) mRNA expression was significantly (p < 0.05) increased in USF + EtOH, but not in SF + EtOH group."
[extra] Obesity (Silver Spring). 2011 Nov;19(11):2280-2. doi: 10.1038/oby.2011.251. Epub 2011 Aug 18. Intestinal permeability is associated with visceral adiposity in healthy women. Gummesson A, Carlsson LM, Storlien LH, Bäckhed F, Lundin P, Löfgren L, Stenlöf K, Lam YY, Fagerberg B, Carlsson B.
[extra] Arterioscler Thromb Vasc Biol. 2013 Jun;33(6):1162-70. doi: 10.1161/ATVBAHA.112.300572. Epub 2013 Apr 4. Dietary carnosine prevents early atherosclerotic lesion formation in apolipoprotein E-null mice. Barski OA, Xie Z, Baba SP, Sithu SD, Agarwal A, Cai J, Bhatnagar A, Srivastava S.
[extra] Br J Pharmacol. 2012 Jun;166(4):1344-56. doi: 10.1111/j.1476-5381.2012.01834.x. D-Carnosine octylester attenuates atherosclerosis and renal disease in ApoE null mice fed a Western diet through reduction of carbonyl stress and inflammation. Menini S, Iacobini C, Ricci C, Scipioni A, Blasetti Fantauzzi C, Giaccari A, Salomone E, Canevotti R, Lapolla A, Orioli M, Aldini G, Pugliese G.
[extra] Can J Physiol Pharmacol. 2004 Jul;82(7):506-14. Amino acid metabolism in the Zucker diabetic fatty rat: effects of insulin resistance and of type 2 diabetes. Wijekoon EP, Skinner C, Brosnan ME, Brosnan JT. "At the insulin-resistant stage, the plasma concentrations of the gluconeogenic amino acids aspartate, serine, glutamine, glycine, and histidine were decreased [...] It is evident that insulin resistance alone is capable of bringing about many of the changes in amino acid metabolism observed in type 2 diabetes."
[extra] Br J Nutr. 2012 Jul 14;108(1):57-61. doi: 10.1017/S0007114511005289. Histidine and arginine are associated with inflammation and oxidative stress in obese women. Niu YC, Feng RN, Hou Y, Li K, Kang Z, Wang J, Sun CH, Li Y. "Among the amino acids determined, serum histidine, arginine, threonine, glycine, lysine and serine were found to be significantly lower in obese women as compared to non-obese controls (P < 0·001). The difference was the greatest for histidine (P < 0·001)."
[extra] Gastroenterology. 2009 Feb;136(2):564-74.e2. doi: 10.1053/j.gastro.2008.09.062. Epub 2008 Oct 7. Dietary histidine ameliorates murine colitis by inhibition of proinflammatory cytokine production from macrophages. Andou A, Hisamatsu T, Okamoto S, Chinen H, Kamada N, Kobayashi T, Hashimoto M, Okutsu T, Shimbo K, Takeda T, Matsumoto H, Sato A, Ohtsu H, Suzuki M, Hibi T.
[extra] Am J Physiol Lung Cell Mol Physiol. 2007 May;292(5):L1095-104. Epub 2007 Jan 12. Protective effect of orally administered carnosine on bleomycin-induced lung injury. Cuzzocrea S, Genovese T, Failla M, Vecchio G, Fruciano M, Mazzon E, Di Paola R, Muià C, La Rosa C, Crimi N, Rizzarelli E, Vancheri C.
[extra] Diabetes. 2013 Jul;62(7):2266-77. doi: 10.2337/db12-1701. Epub 2013 Mar 8. Histidine augments the suppression of hepatic glucose production by central insulin action. Kimura K1, Nakamura Y, Inaba Y, Matsumoto M, Kido Y, Asahara S, Matsuda T, Watanabe H, Maeda A, Inagaki F, Mukai C, Takeda K, Akira S, Ota T, Nakabayashi H, Kaneko S, Kasuga M, Inoue H.
[extra] Hum Exp Toxicol. 2010 Aug;29(8):659-65. The effect of carnosine pretreatment on oxidative stress and hepatotoxicity in binge ethanol administered rats. Artun BC, Küskü-Kiraz Z, Güllüoğlu M, Cevikbaş U, Koçak-Toker N, Uysal M. "In conclusion, carnosine prevented the increases in serum transaminase activities and lipid peroxides in liver of ethanol-treated rats, without any change on steatosis in liver."
[extra] Neurosci Lett. 2012 Feb 21;510(1):1-5. Effects of L-carnosine on splenic sympathetic nerve activity and tumor proliferation. Horii Y, Shen J, Fujisaki Y, Yoshida K, Nagai K. "The tumor volumes of the control mice given water gradually and markedly increased and reached a value of 869.8 ± 132.9 mm3 on day 22 after implantation (Fig. 3b). The tumor volumes of the l-carnosine group also increased; however, the increase was less than that of the control group, reaching a value of 407.8 ± 121.7 mm3 (46.9% of the tumor volume of the control group) on day 22."
[extra] Nutr Clin Pract. 2013 Oct;28(5):609-16. doi: 10.1177/0884533613493333. Epub 2013 Jul 8. Effects of L-carnosine and its zinc complex (Polaprezinc) on pressure ulcer healing. Sakae K, Agata T, Kamide R, Yanagisawa H. "After 4 weeks, the rate of pressure ulcer healing, assessed by the mean weekly improvement in PUSH score, was significantly greater in the CAR (1.6 ± 0.2, P = .02) and PLZ groups (1.8 ± 0.2, P = .009) than in the control group (0.8 ± 0.2)."
[extra] J Periodontol. 2002 Jan;73(1):73-8. Systemic release of endotoxins induced by gentle mastication: association with periodontitis severity. Geerts SO, Nys M, De MP, Charpentier J, Albert A, Legrand V, Rompen EH. "For standardizing the mastication session, the patients were asked to gently chew 2 chewing gums 50 times on each side of the jaw under the supervision of a dental practitioner (SG)." "Overall, blood levels of endotoxin after mastication were found to be significantly higher than before mastication (0.89 +/- 3.3 pg/ml versus 3.0 +/- 5.8 pg/ml; P= 0.0002). Likewise, the incidence of positive endotoxemia rose from 6% before mastication to 24% after mastication (P = 0.001)."
[extra] Arterioscler Thromb Vasc Biol. 2004 Nov;24(11):2174-80. Epub 2004 Sep 23. Severe periodontitis enhances macrophage activation via increased serum lipopolysaccharide. Pussinen PJ, Vilkuna-Rautiainen T, Alfthan G, Palosuo T, Jauhiainen M, Sundvall J, Vesanen M, Mattila K, Asikainen S. "LPS concentration correlated positively with serum concentration of oxidized LDL and cytokine production."
[extra] Innate Immun. 2008 Aug;14(4):247-53. doi: 10.1177/1753425908095130. Lipopolysaccharide associates with pro-atherogenic lipoproteins in periodontitis patients. Kallio KA, Buhlin K, Jauhiainen M, Keva R, Tuomainen AM, Klinge B, Gustafsson A, Pussinen PJ. "Endotoxemia increases with severity of periodontitis. In periodontitis, LPS associates preferentially with the pro-atherogenic VLDL-IDL fraction. Periodontal treatment has only minor effects on plasma LPS activity or distribution, which reflects persistence of the disease."
[extra] Intern Med. 2011;50(15):1569-74. Inflammatory cytokines, adiponectin, insulin resistance and metabolic control after periodontal intervention in patients with type 2 diabetes and chronic periodontitis. Sun WL, Chen LL, Zhang SZ, Wu YM, Ren YZ, Qin GM.
[extra] J Clin Periodontol. 2010 Jan;37(1):53-8. Effect of periodontal treatment on metabolic control, systemic inflammation and cytokines in patients with type 2 diabetes. Correa FO1, Gonçalves D, Figueredo CM, Bastos AS, Gustafsson A, Orrico SR.
[extra] FEBS Letters Volume 581, Issue 3, 6 February 2007, Pages 349–354: Augmentation of 11β-hydroxysteroid dehydrogenase type 1 in LPS-activated J774.1 macrophages – Role of 11β-HSD1 in pro-inflammatory properties in macrophages Takako Ishii, Hiroaki Masuzaki, Tomohiro Tanaka, Naoki Arai, Shintaro Yasue, Nozomi Kobayashi, Tsutomu Tomita, Michio Noguchi, Junji Fujikura, Ken Ebihara, Kiminori Hosoda, Kazuwa Nakao
[extra] Endocrinology. 2011 Apr;152(4):1314-26. doi: 10.1210/en.2010-0659. Epub 2011 Jan 25. Low-grade hypothalamic inflammation leads to defective thermogenesis, insulin resistance, and impaired insulin secretion. Arruda AP, Milanski M, Coope A, Torsoni AS, Ropelle E, Carvalho DP, Carvalheira JB, Velloso LA. "In Wistar rats, hypothalamic TNFα blunts the anorexigenic effect of leptin, which is accompanied by reduced leptin signaling and increased expression of suppressor of cytokine signaling 3. In addition, hypothalamic TNFα reduces O(2) consumption and the expression of thermogenic proteins in brown adipose tissue and skeletal muscle. Furthermore, hypothalamic inflammation increases base-line plasma insulin and insulin secretion by isolated pancreatic islets, which is accompanied by an impaired insulin signal transduction in liver and skeletal muscle. Hypothalamic inflammation induced by stearic acid also reduces O(2) consumption and blunts peripheral insulin signal transduction. The use of intracerebroventricular infliximab restores O(2) consumption in obese rats, whereas TNF receptor 1-knockout mice are protected from diet-induced reduced thermogenesis and defective insulin signal transduction. Thus, low-grade inflammation of the hypothalamus is sufficient to induce changes in a number of parameters commonly impaired in obesity and DM2, and TNFα is an important mediator of this process."
[extra] Diabetes. 2012 Jun;61(6):1455-62. doi: 10.2337/db11-0390. Epub 2012 Apr 20. Inhibition of hypothalamic inflammation reverses diet-induced insulin resistance in the liver. Milanski M, Arruda AP, Coope A, Ignacio-Souza LM, Nunez CE, Roman EA, Romanatto T, Pascoal LB, Caricilli AM, Torsoni MA, Prada PO, Saad MJ, Velloso LA. "Obese rodents were treated by intracerebroventricular injections, with immunoneutralizing antibodies against Toll-like receptor (TLR)4 or tumor necrosis factor (TNF)α, and insulin signal transduction, hepatic steatosis, and gluconeogenesis were evaluated. The inhibition of either TLR4 or TNFα reduced hypothalamic inflammation, which was accompanied by the reduction of hypothalamic resistance to leptin and improved insulin signal transduction in the liver. This was accompanied by reduced liver steatosis and reduced hepatic expression of markers of steatosis. Furthermore, the inhibition of hypothalamic inflammation restored defective liver glucose production"
[extra] J Biol Chem. 2009 Dec 25;284(52):36213-22. doi: 10.1074/jbc.M109.030874. Epub 2009 Oct 26. Deletion of tumor necrosis factor-alpha receptor 1 (TNFR1) protects against diet-induced obesity by means of increased thermogenesis. Romanatto T1, Roman EA, Arruda AP, Denis RG, Solon C, Milanski M, Moraes JC, Bonfleur ML, Degasperi GR, Picardi PK, Hirabara S, Boschero AC, Curi R, Velloso LA.
[extra] Diabetologia. 2007 Jun;50(6):1267-76. Epub 2007 Apr 11. C3H/HeJ mice carrying a toll-like receptor 4 mutation are protected against the development of insulin resistance in white adipose tissue in response to a high-fat diet. Poggi M, Bastelica D, Gual P, Iglesias MA, Gremeaux T, Knauf C, Peiretti F, Verdier M, Juhan-Vague I, Tanti JF, Burcelin R, Alessi MC.
[extra] Obesity (Silver Spring). 2008 Jun;16(6):1248-55. doi: 10.1038/oby.2008.210. Epub 2008 Apr 10. Tlr-4 deficiency selectively protects against obesity induced by diets high in saturated fat. Davis JE1, Gabler NK, Walker-Daniels J, Spurlock ME.
[extra] J Nutr Biochem. 2011 Feb;22(2):136-41. doi: 10.1016/j.jnutbio.2009.12.008. Absence of Tlr2 protects against high-fat diet-induced inflammation and results in greater insulin-stimulated glucose transport in cultured adipocytes. Davis JE, Braucher DR, Walker-Daniels J, Spurlock ME.
[extra] Cell Metab. 2009 Oct;10(4):249-59. doi: 10.1016/j.cmet.2009.08.013. MyD88 signaling in the CNS is required for development of fatty acid-induced leptin resistance and diet-induced obesity. Kleinridders A, Schenten D, Könner AC, Belgardt BF, Mauer J, Okamura T, Wunderlich FT, Medzhitov R, Brüning JC. [Wikipedia: "Model organisms have been used in the study of MYD88 function. The gene was originally discovered and cloned by Dan Liebermann and Barbara Hoffman in mice.[3] In that species it is a universal adapter protein as it is used by almost all TLRs (except TLR 3) to activate the transcription factor NF-κB."]
[extra] Circ Res. 2007 Jun 8;100(11):1589-96. Epub 2007 May 3. Toll-like receptor-4 mediates vascular inflammation and insulin resistance in diet-induced obesity. Kim F1, Pham M, Luttrell I, Bannerman DD, Tupper J, Thaler J, Hawn TR, Raines EW, Schwartz MW.
[extra] FASEB J. 2012 Aug;26(8):3493-502. doi: 10.1096/fj.12-208868. Epub 2012 May 16. Timed high-fat diet resets circadian metabolism and prevents obesity. Sherman H, Genzer Y, Cohen R, Chapnik N, Madar Z, Froy O. "Although timed HF-diet-fed mice consumed the same amount of calories as ad libitum low-fat diet-fed mice, they showed 12% reduced body weight, 21% reduced cholesterol levels, and 1.4-fold increased insulin sensitivity. Compared with the HF diet ad libitum, the timed HF diet led to 18% lower body weight, 30% decreased cholesterol levels, 10% reduced TNF-α levels, and 3.7-fold improved insulin sensitivity. Timed HF-diet-fed mice exhibited a better satiated and less stressed phenotype of 25% lower ghrelin and 53% lower corticosterone levels compared with mice fed the timed low-fat diet."
[extra] FASEB J. 2010 Dec;24(12):4948-59. doi: 10.1096/fj.10-164921. Epub 2010 Aug 19. Germ-free C57BL/6J mice are resistant to high-fat-diet-induced insulin resistance and have altered cholesterol metabolism. Rabot S, Membrez M, Bruneau A, Gérard P, Harach T, Moser M, Raymond F, Mansourian R, Chou CJ.
[extra] PLoS One. 2010 Aug 16;5(8):e12191. doi: 10.1371/journal.pone.0012191. High-fat diet: bacteria interactions promote intestinal inflammation which precedes and correlates with obesity and insulin resistance in mouse. Ding S, Chi MM, Scull BP, Rigby R, Schwerbrock NM, Magness S, Jobin C, Lund PK. "Bacteria and HF diet interact to promote proinflammatory changes in the small intestine, which precede weight gain and obesity and show strong and significant associations with progression of obesity and development of insulin resistance. To our knowledge, this is the first evidence that intestinal inflammation is an early consequence of HF diet which may contribute to obesity and associated insulin resistance. Interventions which limit intestinal inflammation induced by HF diet and bacteria may protect against obesity and insulin resistance."
[extra] Diabetes Care. 2011 Aug;34(8):1809-15. doi: 10.2337/dc10-2197. Epub 2011 Jun 2. Bacterial endotoxin activity in human serum is associated with dyslipidemia, insulin resistance, obesity, and chronic inflammation. Lassenius MI, Pietiläinen KH, Kaartinen K, Pussinen PJ, Syrjänen J, Forsblom C, Pörsti I, Rissanen A, Kaprio J, Mustonen J, Groop PH, Lehto M; FinnDiane Study Group.
[extra] Atherosclerosis. 2009 Apr;203(2):494-502. doi: 10.1016/j.atherosclerosis.2008.06.018. Epub 2008 Jul 1. Ethnic and sex differences in circulating endotoxin levels: A novel marker of atherosclerotic and cardiovascular risk in a British multi-ethnic population. Miller MA, McTernan PG, Harte AL, Silva NF, Strazzullo P, Alberti KG, Kumar S, Cappuccio FP. "Endotoxin levels were positively associated with waist, waist-hip ratio, total cholesterol, serum triglycerides and serum insulin levels and negatively associated with serum HDL-cholesterol."
[extra] Dig Dis Sci. 2012 Jul;57(7):1932-41. doi: 10.1007/s10620-012-2112-9. Epub 2012 Mar 17. Nutrition, intestinal permeability, and blood ethanol levels are altered in patients with nonalcoholic fatty liver disease (NAFLD). Volynets V, Küper MA, Strahl S, Maier IB, Spruss A, Wagnerberger S, Königsrainer A, Bischoff SC, Bergheim I. "Despite no differences in the prevalence of bacterial overgrowth and in the orocecal transit time, intestinal permeability, alcohol, and endotoxin levels in plasma were significantly higher in patients with NAFLD than in controls."
[extra] Surgery. 2012 Apr;151(4):587-93. doi: 10.1016/j.surg.2011.09.038. Epub 2011 Nov 16. Reduction in endotoxemia, oxidative and inflammatory stress, and insulin resistance after Roux-en-Y gastric bypass surgery in patients with morbid obesity and type 2 diabetes mellitus. Monte SV, Caruana JA, Ghanim H, Sia CL, Korzeniewski K, Schentag JJ, Dandona P. "LPS, NF-κB DNA binding, TLR-4, TLR-2, and CD14 expression, CRP, MMP-9, and MCP-1 decreased significantly after RYGB. The mechanism underlying resolution of insulin resistance and T2DM after RYGB may be attributable, at least in part, to the reduction of endotoxemia and associated proinflammatory mediators."
[extra] Diabetes Care. 2009 Dec;32(12):2281-7. doi: 10.2337/dc09-0979. Epub 2009 Sep 15. Increase in plasma endotoxin concentrations and the expression of Toll-like receptors and suppressor of cytokine signaling-3 in mononuclear cells after a high-fat, high-carbohydrate meal: implications for insulin resistance. Ghanim H1, Abuaysheh S, Sia CL, Korzeniewski K, Chaudhuri A, Fernandez-Real JM, Dandona P. "HFHC meal intake induced an increase in plasma LPS concentration and the expression of SOCS-3, TLR2, and TLR4 protein, reactive oxygen species generation, and nuclear factor-kappaB binding activity (P < 0.05 for all). These increases were totally absent after the AHA meal rich in fiber and fruit."
[extra] Diabetologia. 2007 Nov;50(11):2374-83. Epub 2007 Sep 6. Selective increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a mechanism associated with endotoxaemia. Cani PD, Neyrinck AM, Fava F, Knauf C, Burcelin RG, Tuohy KM, Gibson GR, Delzenne NM. "Since bifidobacteria have been reported to reduce intestinal endotoxin levels and improve mucosal barrier function, we specifically increased the gut bifidobacterial content of HF-diet-fed mice through the use of a prebiotic (oligofructose [OFS]). [...] Finally, in HF-OFS-treated-mice, Bifidobacterium spp. significantly and positively correlated with improved glucose tolerance, glucose-induced insulin secretion and normalised inflammatory tone (decreased endotoxaemia, plasma and adipose tissue proinflammatory cytokines)."
[extra] Am J Clin Nutr. 2007 Nov;86(5):1286-92. A high-fat meal induces low-grade endotoxemia: evidence of a novel mechanism of postprandial inflammation. Erridge C, Attina T, Spickett CM, Webb DJ. "Baseline endotoxin concentrations were 8.2 pg/mL (interquartile range: 3.4-13.5 pg/mL) but increased significantly (P < 0.05) by approximately 50% after a high-fat meal or after a high-fat meal with cigarettes but not after no meal or cigarettes alone. These results were validated by the observations that a high-fat meal with or without cigarettes, but not no meal or smoking, also significantly (P < 0.05) reduced plasma endotoxin neutralization capacity, which is an indirect measure of endotoxin exposure"
[extra] Atherosclerosis. 2011 May;216(1):1-6. doi: 10.1016/j.atherosclerosis.2011.02.043. Epub 2011 Feb 26. Diet, commensals and the intestine as sources of pathogen-associated molecular patterns in atherosclerosis, type 2 diabetes and non-alcoholic fatty liver disease. Erridge C. "This review summarises the evidence supporting the proposal that 'pathogen-associated molecular patterns' (PAMPs) derived from dietary and commensal sources may contribute to the chronic inflammatory processes that underpin the development of these diseases via stimulation of TLR2 and TLR4."
[extra] Diabetes Care. 2010 May;33(5):991-7. doi: 10.2337/dc09-1630. Epub 2010 Jan 12. Differential effects of cream, glucose, and orange juice on inflammation, endotoxin, and the expression of Toll-like receptor-4 and suppressor of cytokine signaling-3. Deopurkar R, Ghanim H, Friedman J, Abuaysheh S, Sia CL, Mohanty P, Viswanathan P, Chaudhuri A, Dandona P. "Although both glucose and cream induce NF-kappaB binding and an increase in the expression of SOCS3, TNF-alpha, and IL-1beta in MNCs, only cream caused an increase in LPS concentration and TLR-4 expression. Equicaloric amounts of orange juice or water did not induce a change in any of these indexes. These changes are relevant to the pathogenesis of atherosclerosis and insulin resistance."
[extra] Metabolism. 2006 Sep;55(9):1177-85. Glucose ingestion induces an increase in intranuclear nuclear factor kappaB, a fall in cellular inhibitor kappaB, and an increase in tumor necrosis factor alpha messenger RNA by mononuclear cells in healthy human subjects. Aljada A, Friedman J, Ghanim H, Mohanty P, Hofmeyer D, Chaudhuri A, Dandona P.
[extra] Diabetologia. 2007 Feb;50(2):278-85. Epub 2006 Dec 16. Role of inflammatory mediators in the suppression of insulin receptor phosphorylation in circulating mononuclear cells of obese subjects. Ghanim H, Aljada A, Daoud N, Deopurkar R, Chaudhuri A, Dandona P. "We conclude that in obesity the MNC is characterised by reduced p-INSR-beta and increased inflammatory mediators including IKBKB, PRKCB2 and SOCS3. The increase in SOCS3 but not IKBKB or PRKCB2 is related inversely to p-INSR-beta and might mediate the inhibition of p-INSR-beta."
[extra] Gut. 2012 Dec;61(12):1701-7. doi: 10.1136/gutjnl-2011-301689. Epub 2012 Apr 25. Gut-derived lipopolysaccharide augments adipose macrophage accumulation but is not essential for impaired glucose or insulin tolerance in mice. Caesar R1, Reigstad CS, Bäckhed HK, Reinhardt C, Ketonen M, Lundén GÖ, Cani PD, Bäckhed F.
[extra] J Clin Invest. 2003 Dec;112(12):1796-808. Obesity is associated with macrophage accumulation in adipose tissue. Weisberg SP1, McCann D, Desai M, Rosenbaum M, Leibel RL, Ferrante AW Jr. "Adipose tissue macrophage numbers increase in obesity and participate in inflammatory pathways that are activated in adipose tissues of obese individuals."
[extra] FASEB J. 2008 Jul;22(7):2416-26. doi: 10.1096/fj.07-102723. Epub 2008 Mar 7. Gut microbiota modulation with norfloxacin and ampicillin enhances glucose tolerance in mice. Membrez M, Blancher F, Jaquet M, Bibiloni R, Cani PD, Burcelin RG, Corthesy I, Macé K, Chou CJ.
[extra] PLoS One. 2012;7(3):e32967. doi: 10.1371/journal.pone.0032967. Epub 2012 Mar 7. Leptin resistance in vagal afferent neurons inhibits cholecystokinin signaling and satiation in diet induced obese rats. de Lartigue G, Barbier de la Serre C, Espero E, Lee J, Raybould HE.
[extra] Am J Physiol Endocrinol Metab. 2011 Jul;301(1):E187-95. doi: 10.1152/ajpendo.00056.2011. Epub 2011 Apr 26. Diet-induced obesity leads to the development of leptin resistance in vagal afferent neurons. de Lartigue G, Barbier de la Serre C, Espero E, Lee J, Raybould HE.
[extra] MBio. 2014 Jun 10;5(3). pii: e01011-14. doi: 10.1128/mBio.01011-14. Saccharomyces boulardii Administration Changes Gut Microbiota and Reduces Hepatic Steatosis, Low-Grade Inflammation, and Fat Mass in Obese and Type 2 Diabetic db/db Mice. Everard A, Matamoros S, Geurts L, Delzenne NM, Cani PD.
[extra] Diabetes Metab Syndr Obes. 2012;5:175-89. doi: 10.2147/DMSO.S33473. Epub 2012 Jul 6. Comparison with ancestral diets suggests dense acellular carbohydrates promote an inflammatory microbiota, and may be the primary dietary cause of leptin resistance and obesity. Spreadbury I.
[extra] Gastroenterology. 2012 May;142(5):1100-1101.e2. doi: 10.1053/j.gastro.2012.01.034. Epub 2012 Feb 8. A high-fat diet is associated with endotoxemia that originates from the gut. Pendyala S1, Walker JM, Holt PR. "Placing 8 healthy subjects on a Western-style diet for 1 month induced a 71% increase in plasma levels of endotoxin activity (endotoxemia), whereas a prudent-style diet reduced levels by 31%. The Western-style diet might, therefore, contribute to endotoxemia by causing changes in gastrointestinal barrier function or the composition of the microbiota."
[extra] Cell. 2011 Nov 11;147(4):868-80. doi: 10.1016/j.cell.2011.09.051. CD14 controls the LPS-induced endocytosis of Toll-like receptor 4. Zanoni I, Ostuni R, Marek LR, Barresi S, Barbalat R, Barton GM, Granucci F, Kagan JC. "These data establish that upon microbial detection, an upstream PRR (CD14) controls the trafficking and signaling functions of a downstream PRR (TLR4)."
[extra] Ann Surg. 2003 Feb;237(2):246-55. Protective effects of medium-chain triglycerides on the liver and gut in rats administered endotoxin. Kono H, Fujii H, Asakawa M, Yamamoto M, Matsuda M, Maki A, Matsumoto Y. "All rats given corn oil died after LPS administration; however, this mortality was prevented by MCT in a dose-dependent manner. Rats given corn oil showed liver injury after LPS administration. In contrast, MCT prevented this pathologic change nearly completely. MCT blunted CD14 expression on the Kupffer cells and TNF-alpha production by isolated Kupffer cells; however, there were no differences in phagocytic index between the two groups. The length of the intestinal epithelium was increased in the MCT group compared to the corn oil group. Further, after LPS administration, increases in gut permeability and injury were prevented by MCT. Importantly, MCT also prevented hepatic energy charge and gut injuries in this condition."
[extra] Am J Physiol Gastrointest Liver Physiol. 2013 Dec;305(12):G919-32. doi: 10.1152/ajpgi.00226.2013. Epub 2013 Oct 10. Dietary fat sources differentially modulate intestinal barrier and hepatic inflammation in alcohol-induced liver injury in rats. Zhong W, Li Q, Xie G, Sun X, Tan X, Sun X, Jia W, Zhou Z.
[extra] J Gastroenterol. 2013 Aug 17. Toll-like receptor 4 activation in Barrett's esophagus results in a strong increase in COX-2 expression. Verbeek RE, Siersema PD, Ten Kate FJ, Fluiter K, Souza RF, Vleggaar FP, Bus P, van Baal JW.
[extra] Gastroenterology. 2006 Sep;131(3):862-77. Cox-2 is regulated by Toll-like receptor-4 (TLR4) signaling: Role in proliferation and apoptosis in the intestine. Fukata M, Chen A, Klepper A, Krishnareddy S, Vamadevan AS, Thomas LS, Xu R, Inoue H, Arditi M, Dannenberg AJ, Abreu MT.
[extra] J Leukoc Biol. 2009 Oct;86(4):971-80. doi: 10.1189/jlb.0708396. Epub 2009 Jun 29. TLR4-mediated Cox-2 expression increases intestinal ischemia/reperfusion-induced damage. Moses T, Wagner L, Fleming SD.
[extra] BMC Gastroenterol. 2010 Jul 16;10:82. doi: 10.1186/1471-230X-10-82. The role of prostaglandin E2 (PGE 2) in toll-like receptor 4 (TLR4)-mediated colitis-associated neoplasia. Hernandez Y, Sotolongo J, Breglio K, Conduah D, Chen A, Xu R, Hsu D, Ungaro R, Hayes LA, Pastorini C, Abreu MT, Fukata M.
[extra] Inflamm Bowel Dis. 2011 Jul;17(7):1464-73. Constitutive activation of epithelial TLR4 augments inflammatory responses to mucosal injury and drives colitis-associated tumorigenesis. Fukata M1, Shang L, Santaolalla R, Sotolongo J, Pastorini C, España C, Ungaro R, Harpaz N, Cooper HS, Elson G, Kosco-Vilbois M, Zaias J, Perez MT, Mayer L, Vamadevan AS, Lira SA, Abreu MT.
[extra] J Immunol. 2009 Feb 15;182(4):2476-84. doi: 10.4049/jimmunol.0802059. Lactate boosts TLR4 signaling and NF-kappaB pathway-mediated gene transcription in macrophages via monocarboxylate transporters and MD-2 up-regulation. Samuvel DJ1, Sundararaj KP, Nareika A, Lopes-Virella MF, Huang Y.
[extra] J Neurosci. 2010 Jun 16;30(24):8285-95. doi: 10.1523/JNEUROSCI.0976-10.2010. Pivotal role of TLR4 receptors in alcohol-induced neuroinflammation and brain damage. Alfonso-Loeches S, Pascual-Lucas M, Blanco AM, Sanchez-Vera I, Guerri C.
[extra] Front Cell Neurosci. 2014 May 27;8:146. doi: 10.3389/fncel.2014.00146. eCollection 2014. TLR4-mediated brain inflammation halts neurogenesis: impact of hormonal replacement therapy. Mouihate A. "These data strongly suggest that combined 17β-estradiol and progesterone, and not 17β-estradiol alone, rescues neurogenesis from the deleterious effect of brain inflammation likely via the inhibition of the signaling pathways leading to the activation of proinflammatory genes."
[extra] Wiad Lek. 2013;66(1):3-9. [The role of TLR4 receptor in development of inflammation and carcinogenesis in ulcerative colitis and pharmacotherapy of this disorder]. [Article in Polish] Szumilas D1, Krysiak R, Okopień B. "The increased expression of TLR4 and the development of the uncontrolled inflammatory response in UC (increased production of COX-2, PGE2, TNFalpha and CCL2) impairs regeneration of the mucosa, resulting in its damage, and may later lead to the development of colon cancer."
[extra] AAPS J. 2014 Mar;16(2):246-57. doi: 10.1208/s12248-013-9558-3. Epub 2014 Jan 15. Antitumor activity of gemcitabine can be potentiated in pancreatic cancer through modulation of TLR4/NF-κB signaling by 6-shogaol. Zhou L, Qi L, Jiang L, Zhou P, Ma J, Xu X, Li P. "Overall, our results suggest that 6-shogaol can inhibit the growth of human pancreatic tumors and sensitize them to gemcitabine by suppressing of TLR4/NF-κB-mediated inflammatory pathways linked to tumorigenesis."
[extra] Inflammation. 2014 Apr;37(2):467-77. doi: 10.1007/s10753-013-9760-6. Saturated fatty acids up-regulate COX-2 expression in prostate epithelial cells via toll-like receptor 4/NF-κB signaling. Liu J, Hu S, Cui Y, Sun MK, Xie F, Zhang Q, Jin J.
[extra] Biochem Biophys Res Commun. 2013 Aug 23;438(2):249-56. doi: 10.1016/j.bbrc.2013.07.006. Epub 2013 Jul 11. Differential COX-2 induction by viral and bacterial PAMPs: Consequences for cytokine and interferon responses and implications for anti-viral COX-2 directed therapies. Kirkby NS, Zaiss AK, Wright WR, Jiao J, Chan MV, Warner TD, Herschman HR, Mitchell JA. "LPS induced Cox2 expression in all tissues examined. In contrast, poly(I:C) elicited a milder response, limited to a subset of tissues."
[extra] J Dent Res. 2009 Jun;88(6):519-23. doi: 10.1177/0022034509338353. Is obesity an oral bacterial disease? Goodson JM, Groppo D, Halem S, Carpino E. "Classification tree analysis of salivary microbiological composition revealed that 98.4% of the overweight women could be identified by the presence of a single bacterial species (Selenomonas noxia) at levels greater than 1.05% of the total salivary bacteria."
[extra] Aging Cell. 2013 Dec;12(6):1041-9. doi: 10.1111/acel.12133. Epub 2013 Aug 20. The phytochemical, EGCG, extends lifespan by reducing liver and kidney function damage and improving age-associated inflammation and oxidative stress in healthy rats. Niu Y, Na L, Feng R, Gong L, Zhao Y, Li Q, Li Y, Sun C.
[extra] Biochem Biophys Res Commun. 2012 Oct 5;426(4):480-5. doi: 10.1016/j.bbrc.2012.08.096. Epub 2012 Aug 29. Green tea polyphenol epigallocatechin-3-gallate inhibits TLR4 signaling through the 67-kDa laminin receptor on lipopolysaccharide-stimulated dendritic cells. Byun EB, Choi HG, Sung NY, Byun EH.
[extra] J Immunol. 2010 Jul 1;185(1):33-45. doi: 10.4049/jimmunol.0903742. Epub 2010 May 28. TLR4 signaling inhibitory pathway induced by green tea polyphenol epigallocatechin-3-gallate through 67-kDa laminin receptor. Hong Byun E1, Fujimura Y, Yamada K, Tachibana H.
[extra] Mediators Inflamm. 2009;2009:704706. doi: 10.1155/2009/704706. Epub 2010 Feb 16. Oxymatrine downregulates TLR4, TLR2, MyD88, and NF-kappaB and protects rat brains against focal ischemia. Fan H, Li L, Zhang X, Liu Y, Yang C, Yang Y, Yin J. "The excessive inflammatory immune reaction often resides in region of necrosis and ischemic tissue after cerebral infarction and leads to inflammatory injury. The relationship between toll-like receptor-4 (TLR4), toll-like receptor-2 (TLR2), and nonbiological inflammatory injury has been proved [1–3]. It is believed that injured tissue and necrotic cells release endogenous activators (adjuvants). These activators can combine with TLR4 in the cell membrane. [...] An excessive inflammatory reaction can also damage target cells and tissue [4, 5]. Reducing the inflammatory injury is therefore regarded to be one of major ways to treat acute cerebral infarction."
[extra] Stroke. 2013 Jan;44(1):205-12. Safety and efficacy evaluation of carnosine, an endogenous neuroprotective agent for ischemic stroke. Bae ON, Serfozo K, Baek SH, Lee KY, Dorrance A, Rumbeiha W, Fitzgerald SD, Farooq MU, Naravelta B, Bhatt A, Majid A. "In both permanent and transient ischemic models, carnosine treatment exhibited significant cerebroprotection against histological and functional damage, with wide therapeutic and clinically relevant time windows. Carnosine was well tolerated and exhibited no toxicity."
[extra] Rejuvenation Res. 2008 Feb;11(1):201-14. doi: 10.1089/rej.2007.0608. Dietary supplementation exerts neuroprotective effects in ischemic stroke model. Yasuhara T, Hara K, Maki M, Masuda T, Sanberg CD, Sanberg PR, Bickford PC, Borlongan CV.
[extra] Cardiovasc Res. 2004 Feb 15;61(3):538-47. Modulating Toll-like receptor mediated signaling by (1-->3)-beta-D-glucan rapidly induces cardioprotection. Li C, Ha T, Kelley J, Gao X, Qiu Y, Kao RL, Browder W, Williams DL. "GP treatment reduced infarct size by 47% in rat hearts subjected to reperfusion for 4 h"
[extra] Gut. 2007 Feb;56(2):168-75. Epub 2006 Jun 15. Zinc carnosine, a health food supplement that stabilises small bowel integrity and stimulates gut repair processes. Mahmood A, FitzGerald AJ, Marchbank T, Ntatsaki E, Murray D, Ghosh S, Playford RJ.
[extra] J Clin Biochem Nutr. 2010 May;46(3):234-43. doi: 10.3164/jcbn.09-125. Epub 2010 Apr 10. Polaprezinc Protects Mice against Endotoxin Shock. Ohata S, Moriyama C, Yamashita A, Nishida T, Kusumoto C, Mochida S, Minami Y, Nakada J, Shomori K, Inagaki Y, Ohta Y, Matsura T. "PZ pretreatment inhibited the decrease in the survival rate of mice after LPS injection. PZ inhibited the increases in plasma NO as well as TNF-alpha after LPS. Compatibly, PZ suppressed LPS-induced inducible NO synthase mRNA transcription in the mouse lungs. PZ also improved LPS-induced lung injury."
[extra] ISME J. 2013 Apr;7(4):880-4. doi: 10.1038/ismej.2012.153. Epub 2012 Dec 13. An opportunistic pathogen isolated from the gut of an obese human causes obesity in germfree mice. Fei N, Zhao L.
[extra] J Am Coll Cardiol. 1999 Dec;34(7):1975-81. Association of endotoxemia with carotid atherosclerosis and cardiovascular disease: prospective results from the Bruneck Study. Wiedermann CJ1, Kiechl S, Dunzendorfer S, Schratzberger P, Egger G, Oberhollenzer F, Willeit J. "Median endotoxin concentration amounted to 14.3 pg/ml (range, 6.0 to 209.2 pg/ml). Subjects with levels beyond 50 pg/ml (90th percentile) faced a threefold risk of incident atherosclerosis (odds ratio [95% confidence interval] 2.9 [1.4-6.3]; p < 0.01). The risk associated with high endotoxin was most pronounced in subjects with chronic infections and in current and ex-smokers."
[extra] Arterioscler Thromb Vasc Biol. 2007 Jun;27(6):1433-9. Epub 2007 Mar 15. Endotoxemia, immune response to periodontal pathogens, and systemic inflammation associate with incident cardiovascular disease events. Pussinen PJ, Tuomisto K, Jousilahti P, Havulinna AS, Sundvall J, Salomaa V. "The corresponding HRs for high endotoxin concentration were 1.82 (1.22 to 2.73, alone), 3.92 (1.99 to 7.74, with CRP), 3.54 (1.78 to 7.03, with IL-6), and 2.26 (1.13 to 4.52, with tumor necrosis factor (TNF)-alpha) after adjusting for age and gender. These associations were abolished after adjusting for serum lipids."
[extra] Arterioscler Thromb Vasc Biol. 2004 Dec;24(12):2227-36. Epub 2004 Oct 7. Potential role of endotoxin as a proinflammatory mediator of atherosclerosis. Stoll LL, Denning GM, Weintraub NL. "In this article, we outline the main elements of the endotoxin signaling receptor complex that initiates proinflammatory signaling (lipopolysaccharide binding protein [LBP], CD14, TLR-4, and MD-2) and discuss how changes in expression of these molecules may affect proatherogenic responses in the vessel wall. We also describe some of the proinflammatory effects of endotoxin that may be relevant to atherosclerosis, and discuss how serum lipoproteins, especially high-density lipoprotein, may modulate endotoxin-induced inflammatory responses. Further, we discuss recent findings suggesting that the lipid-lowering statins may have an additional protective role in blocking at least some of these proinflammatory signaling pathways."
[extra] Metab Brain Dis. 2014 Mar;29(1):19-36. doi: 10.1007/s11011-013-9435-x. Epub 2013 Sep 10. Mitochondrial dysfunctions in myalgic encephalomyelitis/chronic fatigue syndrome explained by activated immuno-inflammatory, oxidative and nitrosative stress pathways. Morris G, Maes M. "Increased levels of pro-inflammatory cytokines, such as interleukin-1 and tumor necrosis factor-α, and elastase, and increased O&NS may inhibit mitochondrial respiration, decrease the activities of the electron transport chain and mitochondrial membrane potential, increase mitochondrial membrane permeability, interfere with ATP production and cause mitochondrial shutdown. The activated O&NS pathways may additionally lead to damage of mitochondrial DNA and membranes thus decreasing membrane fluidity. Lowered levels of antioxidants, zinc and coenzyme Q10, and ω3 polyunsaturated fatty acids in ME/cfs may further aggravate the activated immuno-inflammatory and O&NS pathways."hi
[extra] Neurochem Res. 2005 Oct;30(10):1283-8. Carnosine protects the brain of rats and Mongolian gerbils against ischemic injury: after-stroke-effect. Dobrota D, Fedorova T, Stvolinsky S, Babusikova E, Likavcanova K, Drgova A, Strapkova A, Boldyrev A. "In this study, the therapeutic effect of carnosine was estimated on animals when this drug was administered intraperitoneally (100 mg/kg body weight) after ischemic episode induced by experimental global brain ischemia. Treatment of the animals with carnosine after ischemic episode under long-term (7-14 days) reperfusion demonstrated its pronounced protective effect on neurological symptoms and animal mortality. Carnosine also prevented higher lipid peroxidation of brain membrane structures and increased a resistance of neuronal membranes to the in vitro induced oxidation. Measurements of malonyl dialdehyde (MDA) in brain homogenates showed its increase in the after brain stroke animals and decreased MDA level in the after brain stroke animals treated with carnosine."
[extra] Arch Gerontol Geriatr. 2014 May 2. Anserine and carnosine supplementation in the elderly: Effects on cognitive functioning and physical capacity. Szcześniak D, Budzeń S, Kopeć W, Rymaszewska J. [13 weeks, 0.33g/d carnosine, 0.66g/d anserine -> BMI decreased by 0.51]
[extra] Journal of Human Kinetics, Volume 40, Issue 1 (Mar 2014). Effect of Choline Supplementation on Rapid Weight Loss and Biochemical Variables Among Female Taekwondo and Judo Athlete. Gehan Elsawy, Osama Abdelrahman, Amr Hamza. [significantly increased fat loss and decreased MDA with 4g/d choline]
[extra] Diabetes. 2006 Sep;55(9):2554-61. Exercise improves insulin and leptin sensitivity in hypothalamus of Wistar rats. Flores MB, Fernandes MF, Ropelle ER, Faria MC, Ueno M, Velloso LA, Saad MJ, Carvalheira JB.
[extra] PLoS Biol. 2010 Aug 24;8(8). pii: e1000465. doi: 10.1371/journal.pbio.1000465. IL-6 and IL-10 anti-inflammatory activity links exercise to hypothalamic insulin and leptin sensitivity through IKKbeta and ER stress inhibition. Ropelle ER, Flores MB, Cintra DE, Rocha GZ, Pauli JR, Morari J, de Souza CT, Moraes JC, Prada PO, Guadagnini D, Marin RM, Oliveira AG, Augusto TM, Carvalho HF, Velloso LA, Saad MJ, Carvalheira JB.
[extra] Mol Cell Biol. 2014 Jul 28. pii: MCB.00157-14. [Epub ahead of print] 12-Lipoxygenase Promotes Obesity-Induced Oxidative Stress in Pancreatic Islets. Tersey SA, Maier B, Nishiki Y, Maganti AV, Nadler JL, Mirmira RG. "High fat diets lead to obesity, inflammation, and dysglycemia. 12-lipoxygenase (12-LO) is activated by high fat diets and catalyzes the oxygenation of cellular arachidonic acid to form proinflammatory intermediates. [...] Whereas knockout mice and control littermates displayed identical weight gain, body fat distribution, and macrophage infiltration into fat, knockout mice exhibited greater adaptive islet hyperplasia, improved insulin secretion, and complete protection from dysglycemia."
[extra] Mol Cell Biochem. 2009 Feb;322(1-2):127-35. doi: 10.1007/s11010-008-9949-4. Epub 2008 Nov 11. Curcumin attenuates inflammation through inhibition of TLR-4 receptor in experimental colitis. Lubbad A, Oriowo MA, Khan I. "TNBS-induced increase in the level of MPO activity and MDA concentrations was reversed by curcumin treatment, whereas the same dose of curcumin did not affect their levels in the non-colitis animals. Increases in the levels of TLR-4, MyD88, and NFkB proteins in inflamed tissue were also suppressed significantly by curcumin treatment."
[extra] Cell. 2014 Aug 14;158(4):705-21. doi: 10.1016/j.cell.2014.05.052. Altering the Intestinal Microbiota during a Critical Developmental Window Has Lasting Metabolic Consequences. Cox LM, Yamanishi S, Sohn J, Alekseyenko AV, Leung JM, Cho I, Kim SG, Li H, Gao Z, Mahana D, Zárate Rodriguez JG, Rogers AB, Robine N, Loke P, Blaser MJ. "LDP that is limited to early life transiently perturbs the microbiota, which is sufficient to induce sustained effects on body composition, indicating that microbiota interactions in infancy may be critical determinants of long-term host metabolic effects. In addition, LDP enhances the effect of high-fat diet induced obesity. The growth promotion phenotype is transferrable to germ-free hosts by LDP-selected microbiota, showing that the altered microbiota, not antibiotics per se, play a causal role."
[extra] Diabetes Care. 2008 Sep;31(9):1827-31. doi: 10.2337/dc08-0561. Epub 2008 Jun 12. Acute modulation of toll-like receptors by insulin. Ghanim H, Mohanty P, Deopurkar R, Sia CL, Korzeniewski K, Abuaysheh S, Chaudhuri A, Dandona P.
[extra] Diabetes Care. 2010 Nov;33(11):2416-23. doi: 10.2337/dc10-0929. Epub 2010 Aug 10. Insulin suppresses endotoxin-induced oxidative, nitrosative, and inflammatory stress in humans. Dandona P, Ghanim H, Bandyopadhyay A, Korzeniewski K, Ling Sia C, Dhindsa S, Chaudhuri A. "The coinfusion of insulin led to a total elimination of the increase in NOM, free fatty acids, and TBARS and a significant reduction in ROS generation by PMNLs and plasma MIF, visfatin, and myoglobin concentrations. Insulin did not affect TNF-α, MCP-1, IL-6, LBP, resistin, and HMG-B1 increases induced by the LPS."
[extra] Oncogene. 2014 Jan 2;33(1):74-84. doi: 10.1038/onc.2012.540. Epub 2012 Nov 26. Enhancement of leptin receptor signaling by SOCS3 deficiency induces development of gastric tumors in mice. Inagaki-Ohara K, Mayuzumi H, Kato S, Minokoshi Y, Otsubo T, Kawamura YI, Dohi T, Matsuzaki G, Yoshimura A. "These data suggest that SOCS3 is an antigastric tumor gene that suppresses leptin overexpression and ObRb/STAT3 hyperactivation, supporting the hypothesis that the leptin/ObRb/STAT3 axis accelerates tumorigenesis and that it may represent a new therapeutic target for the treatment of gastric cancer."
[extra] Nat Med. 2004 Jul;10(7):734-8. Epub 2004 Jun 27. Enhanced leptin sensitivity and attenuation of diet-induced obesity in mice with haploinsufficiency of Socs3. Howard JK, Cave BJ, Oksanen LJ, Tzameli I, Bjørbaek C, Flier JS.
[extra] Nat Med. 2004 Jul;10(7):739-43. Epub 2004 Jun 20. Socs3 deficiency in the brain elevates leptin sensitivity and confers resistance to diet-induced obesity. Mori H, Hanada R, Hanada T, Aki D, Mashima R, Nishinakamura H, Torisu T, Chien KR, Yasukawa H, Yoshimura A. "These data indicate that Socs3 is a key regulator of diet-induced leptin as well as insulin resistance. Our study demonstrates the negative regulatory role of Socs3 in leptin signaling in vivo, and thus suppression of Socs3 in the brain is a potential therapy for leptin-resistance in obesity."
[extra] Dev Cell. 2002 Apr;2(4):489-95. PTP1B regulates leptin signal transduction in vivo. Zabolotny JM, Bence-Hanulec KK, Stricker-Krongrad A, Haj F, Wang Y, Minokoshi Y, Kim YB, Elmquist JK, Tartaglia LA, Kahn BB, Neel BG.
[extra] Am J Physiol Endocrinol Metab. 2013 Sep 1;305(5):E649-59. Acute exercise suppresses hypothalamic PTP1B protein level and improves insulin and leptin signaling in obese rats. Chiarreotto-Ropelle EC, Pauli LS, Katashima CK, Pimentel GD, Picardi PK, Silva VR, de Souza CT, Prada PO, Cintra DE, Carvalheira JB, Ropelle ER, Pauli JR. "Interestingly, the anti-inflammatory action and the reduction of PTP1B activity mediated by exercise occurred in an interleukin-6 (IL-6)-dependent manner because exercise failed to reduce inflammation and PTP1B protein level after the disruption of hypothalamic-specific IL-6 action in obese rats. Conversely, intracerebroventricular administration of recombinant IL-6 reproduced the effects of exercise, improving hypothalamic insulin and leptin action by reducing the inflammatory signaling and PTP1B activity in obese rats at rest. Taken together, our study reports that physical exercise restores insulin and leptin signaling, at least in part, by reducing hypothalamic PTP1B protein level through the central anti-inflammatory response."
[extra] Nat Med. 2006 Aug;12(8):917-24. Epub 2006 Jul 16. Neuronal PTP1B regulates body weight, adiposity and leptin action. Bence KK, Delibegovic M, Xue B, Gorgun CZ, Hotamisligil GS, Neel BG, Kahn BB. "Thus, PTP1B regulates body mass and adiposity primarily through actions in the brain. Furthermore, neuronal PTP1B regulates adipocyte leptin production and probably is essential for the development of leptin resistance."
[extra] Cell. 2008 Oct 3;135(1):61-73. doi: 10.1016/j.cell.2008.07.043. Hypothalamic IKKbeta/NF-kappaB and ER stress link overnutrition to energy imbalance and obesity. Zhang X, Zhang G, Zhang H, Karin M, Bai H, Cai D. "Overnutrition atypically activates hypothalamic IKKbeta/NF-kappaB at least in part through elevated endoplasmic reticulum stress in the hypothalamus. While forced activation of hypothalamic IKKbeta/NF-kappaB interrupts central insulin/leptin signaling and actions, site- or cell-specific suppression of IKKbeta either broadly across the brain or locally within the mediobasal hypothalamus, or specifically in hypothalamic AGRP neurons significantly protects against obesity and glucose intolerance. The molecular mechanisms involved include regulation by IKKbeta/NF-kappaB of SOCS3, a core inhibitor of insulin and leptin signaling."
[extra] J Clin Invest. 2009 Sep;119(9):2577-89. doi: 10.1172/JCI36714. Epub 2009 Aug 10. Palmitic acid mediates hypothalamic insulin resistance by altering PKC-theta subcellular localization in rodents. Benoit SC, Kemp CJ, Elias CF, Abplanalp W, Herman JP, Migrenne S, Lefevre AL, Cruciani-Guglielmacci C, Magnan C, Yu F, Niswender K, Irani BG, Holland WL, Clegg DJ. "CNS exposure to palmitic acid via direct infusion or by oral gavage increased the localization of PKC-theta to cell membranes in the hypothalamus, which was associated with impaired hypothalamic insulin and leptin signaling. This finding was specific for palmitic acid, as the monounsaturated fatty acid, oleic acid, neither increased membrane localization of PKC-theta nor induced insulin resistance. Finally, arcuate-specific knockdown of PKC-theta attenuated diet-induced obesity and improved insulin signaling. These results suggest that many of the deleterious effects of high-fat diets, specifically those enriched with palmitic acid, are CNS mediated via PKC-theta activation, resulting in reduced insulin activity."
[extra] Diabetes. 1999 Jun;48(6):1270-4. Free fatty acid-induced insulin resistance is associated with activation of protein kinase C theta and alterations in the insulin signaling cascade. Griffin ME, Marcucci MJ, Cline GW, Bell K, Barucci N, Lee D, Goodyear LJ, Kraegen EW, White MF, Shulman GI. "We conclude that acute elevations of plasma FFA levels for 5 h induce skeletal muscle insulin resistance in vivo via a reduction in insulin-stimulated muscle glycogen synthesis and glucose oxidation that can be attributed to reduced glucose transport activity. These changes are associated with abnormalities in the insulin signaling cascade and may be mediated by FFA activation of PKC theta."
[extra] Mol Immunol. 2009 Dec;47(2-3):420-5. doi: 10.1016/j.molimm.2009.08.023. Epub 2009 Sep 20. Molecular mechanism underlying the suppression of lipid oxidation during endotoxemia. Maitra U, Chang S, Singh N, Li L. "In this report, we have demonstrated that LPS suppresses free fatty acid (FFA) oxidation, and consequently contributes to elevated plasma levels of FFA and triglyceride (TG). Furthermore, this process depends upon the interleukin-1 receptor associated kinase 1 (IRAK-1), one of the key TLR4 intracellular signaling kinases. IRAK-1(-/-) mice fail to exhibit the dramatic rise in plasma FFA and TG levels compared to wild-type (WT) mice following lethal LPS injection. Mechanistically, we demonstrated that LPS suppresses FFA oxidation through decreasing the expression levels of key FFA oxidative genes including CPT-1 and MCAD in both liver and kidney tissues of WT but not IRAK-1(-/-) mice."
[extra] Mol Immunol. 2012 Apr;50(4):244-52. Reduced oxidative tissue damage during endotoxemia in IRAK-1 deficient mice. Singh N, Li L. "In contrast, LPS challenge failed to reduce the activities of GPx and SOD in IRAK-1 deficient tissues."
[extra] Chang, Samantha, and Liwu Li. "Metabolic endotoxemia: a novel concept in chronic disease pathology." J Med Sci 3 (2011): 191-209.
[extra] Hepatology. 2003 Mar;37(3):551-7. Oral bile acids reduce bacterial overgrowth, bacterial translocation, and endotoxemia in cirrhotic rats. Lorenzo-Zúñiga V, Bartolí R, Planas R, Hofmann AF, Viñado B, Hagey LR, Hernández JM, Mañé J, Alvarez MA, Ausina V, Gassull MA.
[extra] Proc Natl Acad Sci U S A. 2004 Nov 2;101(44):15718-23. The gut microbiota as an environmental factor that regulates fat storage. Bäckhed F, Ding H, Wang T, Hooper LV, Koh GY, Nagy A, Semenkovich CF, Gordon JI. "We found that conventionalization of adult germ-free (GF) C57BL/6 mice with a normal microbiota harvested from the distal intestine (cecum) of conventionally raised animals produces a 60% increase in body fat content and insulin resistance within 14 days despite reduced food intake."
[extra] ISME J. 2013 Apr;7(4):880-4. doi: 10.1038/ismej.2012.153. Epub 2012 Dec 13. An opportunistic pathogen isolated from the gut of an obese human causes obesity in germfree mice. Fei N, Zhao L."Here we show that one endotoxin-producing bacterium isolated from a morbidly obese human's gut induced obesity and insulin resistance in germfree mice. The endotoxin-producing Enterobacter decreased in relative abundance from 35% of the volunteer's gut bacteria to non-detectable, during which time the volunteer lost 51.4 kg of 174.8 kg initial weight and recovered from hyperglycemia and hypertension after 23 weeks on a diet of whole grains, traditional Chinese medicinal foods and prebiotics. [...] Mono-association of germfree C57BL/6J mice with strain Enterobacter cloacae B29 isolated from the volunteer's gut induced fully developed obesity and insulin resistance on a high-fat diet but not on normal chow diet, whereas the germfree control mice on a high-fat diet did not exhibit the same disease phenotypes."
[extra] Gastroenterology. 2012 Oct;143(4):913-6.e7. Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Vrieze A, Van Nood E, Holleman F, Salojärvi J, Kootte RS, Bartelsman JF, Dallinga-Thie GM, Ackermans MT, Serlie MJ, Oozeer R, Derrien M, Druesne A, Van Hylckama Vlieg JE, Bloks VW, Groen AK, Heilig HG, Zoetendal EG, Stroes ES, de Vos WM, Hoekstra JB, Nieuwdorp M. "Subjects were assigned randomly to groups that were given small intestinal infusions of allogenic or autologous microbiota. Six weeks after infusion of microbiota from lean donors, insulin sensitivity of recipients increased (median rate of glucose disappearance changed from 26.2 to 45.3 μmol/kg/min; P < .05) along with levels of butyrate-producing intestinal microbiota."
[21] Am J Clin Nutr. 2010 Apr;91(4):940-9. Orange juice neutralizes the proinflammatory effect of a high-fat, high-carbohydrate meal and prevents endotoxin increase and Toll-like receptor expression. Ghanim H, Sia CL, Upadhyay M, Korzeniewski K, Viswanathan P, Abuaysheh S, Mohanty P, Dandona P.
[22] J Clin Endocrinol Metab. 2011 May;96(5):1409-14. A resveratrol and polyphenol preparation suppresses oxidative and inflammatory stress response to a high-fat, high-carbohydrate meal. Ghanim H, Sia CL, Korzeniewski K, Lohano T, Abuaysheh S, Marumganti A, Chaudhuri A, Dandona P.
[23] Endocrine Abstracts (2012) The intake of fiber suppresses the high fat high carbohydrate meal induced endotoxemia, oxidative stress and inflammation H. Ghanim, M. Batra, S. Dhindsa, K. Green, S. Abuaysheh, A. Chaudhuri & P. Dandona
[24] Br J Nutr. 2009 Apr;101(7):961-6. Probiotic yogurt in the elderly with intestinal bacterial overgrowth: endotoxaemia and innate immune functions. Schiffrin EJ1, Parlesak A, Bode C, Bode JC, van't Hof MA, Grathwohl D, Guigoz Y.
[25] Nutrition. 2012 May;28(5):539-43. Probiotic yogurt improves antioxidant status in type 2 diabetic patients. Ejtahed HS1, Mohtadi-Nia J, Homayouni-Rad A, Niafar M, Asghari-Jafarabadi M, Mofid V. "Probiotic yogurt significantly decreased fasting blood glucose (P < 0.01) and hemoglobin A1c (P < 0.05) and increased erythrocyte superoxide dismutase and glutathione peroxidase activities and total antioxidant status (P < 0.05) compared with the control group [conventional yogurt]."
[26] Ann Nutr Metab. 2013;63(1-2):1-9. Effect of multispecies probiotic supplements on metabolic profiles, hs-CRP, and oxidative stress in patients with type 2 diabetes. Asemi Z1, Zare Z, Shakeri H, Sabihi SS, Esmaillzadeh A. "prevented a rise in FPG and resulted in a decrease in serum hs-CRP and an increase in plasma total GSH"
[27] Iran J Pharm Res. 2013 Fall;12(4):929-36. Probiotic yogurt Affects Pro- and Anti-inflammatory Factors in Patients with Inflammatory Bowel Disease. Shadnoush M, Shaker Hosseini R, Mehrabi Y, Delpisheh A, Alipoor E, Faghfoori Z, Mohammadpour N, Zaringhalam Moghadam J.
[28] Biochem Biophys Res Commun. 2013 Feb 8;431(2):258-63. Lactobacillus rhamnosus GG improves insulin sensitivity and reduces adiposity in high-fat diet-fed mice through enhancement of adiponectin production. Kim SW, Park KY, Kim B, Kim E, Hyun CK.
[29] PLoS One. 2014 Jan 27;9(1):e80169. Lactobacillus rhamnosus GG protects against non-alcoholic fatty liver disease in mice. Ritze Y1, Bárdos G1, Claus A1, Ehrmann V1, Bergheim I2, Schwiertz A3, Bischoff SC1.
[30] Am J Pathol. 2011 Dec;179(6):2866-75. Lactobacillus rhamnosus GG treatment potentiates intestinal hypoxia-inducible factor, promotes intestinal integrity and ameliorates alcohol-induced liver injury. Wang Y1, Kirpich I, Liu Y, Ma Z, Barve S, McClain CJ, Feng W.
[31] J Nutr Biochem. 2013 Sep;24(9):1609-15. Lactobacillus rhamnosus GG reduces hepatic TNFα production and inflammation in chronic alcohol-induced liver injury. Wang Y1, Liu Y, Kirpich I, Ma Z, Wang C, Zhang M, Suttles J, McClain C, Feng W.
[32] World J Gastroenterol. 2013 Jan 14;19(2):274-83. Dual probiotic strains suppress high fructose-induced metabolic syndrome. Park DY, Ahn YT, Huh CS, McGregor RA, Choi MS.
[33] Diabetologia. 2013 May;56(5):985-94. Histidine supplementation improves insulin resistance through suppressed inflammation in obese women with the metabolic syndrome: a randomised controlled trial. Feng RN, Niu YC, Sun XW, Li Q, Zhao C, Wang C, Guo FC, Sun CH, Li Y.
[34] Eur J Pharmacol. 2011 Feb 25;653(1-3):82-8. Histidine and carnosine alleviated hepatic steatosis in mice consumed high saturated fat diet. Mong MC1, Chao CY, Yin MC.
[35] Eur J Pharmacol. 2005 Apr 18;513(1-2):145-50. Histidine and carnosine delay diabetic deterioration in mice and protect human low density lipoprotein against oxidation and glycation. Lee YT, Hsu CC, Lin MH, Liu KS, Yin MC. "1 g/l histidine and carnosine treatments significantly reduced cholesterol level in heart and liver (P < 0.05). The administration of histidine or carnosine significantly enhanced catalase activity and decreased lipid oxidation levels in kidney and liver (P < 0.05); however, only 1 g/l histidine and carnosine treatments significantly increased glutathione peroxidase activity (P < 0.05). The increased interleukin (IL)-6 and tumor necrosis factor (TNF)-alpha in diabetic mice were significantly suppressed by the intake of histidine or carnosine (P < 0.05)."
[36] Food Chem Toxicol. 2008 May;46(5):1503-9. Beneficial effects of histidine and carnosine on ethanol-induced chronic liver injury. Liu WH, Liu TC, Yin MC.
[37] J Food Sci. 2009 Oct;74(8):H259-65. Protective effects from carnosine and histidine on acetaminophen-induced liver injury. Yan SL, Wu ST, Yin MC, Chen HT, Chen HC.
[38] Tohoku J Exp Med. 2000 Jun;191(2):85-99. Effect of carnosine on rats under experimental brain ischemia. Gallant S, Kukley M, Stvolinsky S, Bulygina E, Boldyrev A.
[39] Nutr Res Pract. 2011 Oct;5(5):421-8. Effects of α-lipoic acid and L-carnosine supplementation on antioxidant activities and lipid profiles in rats. Kim MY1, Kim EJ, Kim YN, Choi C, Lee BH.
[40] PLoS One. 2011 Mar 15;6(3):e17971. Effects of dietary supplementation of carnosine on mitochondrial dysfunction, amyloid pathology, and cognitive deficits in 3xTg-AD mice. Corona C, Frazzini V, Silvestri E, Lattanzio R, La Sorda R, Piantelli M, Canzoniero LM, Ciavardelli D, Rizzarelli E, Sensi SL.
[41] J Physiol Biochem. 2014 Jun;70(2):385-95. Effect of carnosine alone or combined with α-tocopherol on hepatic steatosis and oxidative stress in fructose-induced insulin-resistant rats. Giriş M, Doğru-Abbasoğlu S, Kumral A, Olgaç V, Koçak-Toker N, Uysal M.
[42] Diabetes. 2009 Jul;58(7):1509-17. doi: 10.2337/db08-1637. Epub 2009 Apr 14. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Gao Z, Yin J, Zhang J, Ward RE, Martin RJ, Lefevre M, Cefalu WT, Ye J. "On the high-fat diet, supplementation of butyrate prevented development of insulin resistance and obesity in C57BL/6 mice."
[43] Am J Pathol. 2004 May;164(5):1547-56. Abrogation of nuclear factor-kappaB activation is involved in zinc inhibition of lipopolysaccharide-induced tumor necrosis factor-alpha production and liver injury. Zhou Z, Wang L, Song Z, Saari JT, McClain CJ, Kang YJ.
[44] J Pharmacol Exp Ther. 2003 Jun;305(3):880-6. Epub 2003 Mar 6. Prevention of alterations in intestinal permeability is involved in zinc inhibition of acute ethanol-induced liver damage in mice. Lambert JC, Zhou Z, Wang L, Song Z, McClain CJ, Kang YJ.
[45] Proc Natl Acad Sci U S A. 2013 Apr 23;110(17):7003-8. Intestinal alkaline phosphatase prevents metabolic syndrome in mice. Kaliannan K, Hamarneh SR, Economopoulos KP, Nasrin Alam S, Moaven O, Patel P, Malo NS, Ray M, Abtahi SM, Muhammad N, Raychowdhury A, Teshager A, Mohamed MM, Moss AK, Ahmed R, Hakimian S, Narisawa S, Millán JL, Hohmann E, Warren HS, Bhan AK, Malo MS, Hodin RA.
[46] J Nutr Sci Vitaminol (Tokyo). 2007 Jun;53(3):219-24. Enhancement effects of vitamin K1 (phylloquinone) or vitamin K2 (menaquinone-4) on intestinal alkaline phosphatase activity in rats. Sogabe N1, Maruyama R, Hosori T, Goseki-Sone M.
[47] Bone. 2011 May 1;48(5):1036-42. Effects of long-term vitamin K(1) (phylloquinone) or vitamin K(2) (menaquinone-4) supplementation on body composition and serum parameters in rats. Sogabe N1, Maruyama R, Baba O, Hosoi T, Goseki-Sone M.
[48] Int J Environ Res Public Health. 2009 Oct;6(10):2626-38. Food choices and coronary heart disease: a population based cohort study of rural Swedish men with 12 years of follow-up. Holmberg S, Thelin A, Stiernström EL.
[49] J Neurochem. 2007 May;101(3):729-36. Epub 2007 Jan 24. Neuroprotective actions of a histidine analogue in models of ischemic stroke. Tang SC, Arumugam TV, Cutler RG, Jo DG, Magnus T, Chan SL, Mughal MR, Telljohann RS, Nassar M, Ouyang X, Calderan A, Ruzza P, Guiotto A, Mattson MP. "We recently synthesized and characterized histidine analogues related to the natural dipeptide carnosine, which selectively scavenge the toxic lipid peroxidation product 4-hydroxynonenal (HNE)[...]"
[50] J Clin Endocrinol Metab. 2007 Nov;92(11):4180-4. Epub 2007 Aug 14. Association between hypothyroidism and small intestinal bacterial overgrowth. Lauritano EC, Bilotta AL, Gabrielli M, Scarpellini E, Lupascu A, Laginestra A, Novi M, Sottili S, Serricchio M, Cammarota G, Gasbarrini G, Pontecorvi A, Gasbarrini A.
[51] Ann Rheum Dis. 2004 Apr;63(4):450-2. A link between irritable bowel syndrome and fibromyalgia may be related to findings on lactulose breath testing. Pimentel M, Wallace D, Hallegua D, Chow E, Kong Y, Park S, Lin HC.
[52] Am J Gastroenterol. 2002 Sep;97(9):2364-70. Small intestinal bacterial overgrowth in human cirrhosis is associated with systemic endotoxemia. Bauer TM, Schwacha H, Steinbrückner B, Brinkmann FE, Ditzen AK, Aponte JJ, Pelz K, Berger D, Kist M, Blum HE.
[53] Br J Nutr. 2012 Jan;107(2):229-41. doi: 10.1017/S0007114511002868. Epub 2011 Jun 29. Saturated fatty acids activate microglia via Toll-like receptor 4/NF-κB signalling. Wang Z, Liu D, Wang F, Liu S, Zhao S, Ling EA, Hao A.
[54] J Biol Chem. 2001 May 18;276(20):16683-9. Epub 2001 Mar 2. Saturated fatty acids, but not unsaturated fatty acids, induce the expression of cyclooxygenase-2 mediated through Toll-like receptor 4. Lee JY, Sohn KH, Rhee SH, Hwang D.
[55] Arterioscler Thromb Vasc Biol. 2007 Jan;27(1):84-91. Epub 2006 Nov 2. Role of the Toll-like receptor 4/NF-kappaB pathway in saturated fatty acid-induced inflammatory changes in the interaction between adipocytes and macrophages. Suganami T, Tanimoto-Koyama K, Nishida J, Itoh M, Yuan X, Mizuarai S, Kotani H, Yamaoka S, Miyake K, Aoe S, Kamei Y, Ogawa Y. "These findings suggest that saturated FAs, which are released in large quantities from hypertrophied adipocytes via the macrophage-induced adipocyte lipolysis, serve as a naturally occurring ligand for TLR4, thereby inducing the inflammatory changes in both adipocytes and macrophages through NF-kappaB activation."
[56] J Neurochem. 2012 Mar;120(6):1060-71. doi: 10.1111/j.1471-4159.2012.07660.x. Epub 2012 Feb 6. Saturated long-chain fatty acids activate inflammatory signaling in astrocytes. Gupta S, Knight AG, Gupta S, Keller JN, Bruce-Keller AJ. "Data show that the saturated fatty acid palmitic acid, as well as lauric acid and stearic acid, trigger the release of TNFα and IL-6 from astrocytes. Unsaturated fatty acids were unable to induce cytokine release from cultured astrocytes. Furthermore, the effects of palmitic acid on cytokine release require Toll-like receptor 4 rather than CD36 or Toll-like receptor 2, and do not depend on palmitic acid metabolism to palmitoyl-CoA. [...] Finally, data show that the essential ω-3 fatty acid docosahexaenoic acid acts in a dose-dependent manner to prevent the actions of palmitic acid on inflammatory signaling in astrocytes. Collectively, these data demonstrate the ability of saturated fatty acids to induce astrocyte inflammation in vitro."
[57] J Neurosci. 2009 Jan 14;29(2):359-70. doi: 10.1523/JNEUROSCI.2760-08.2009. Saturated fatty acids produce an inflammatory response predominantly through the activation of TLR4 signaling in hypothalamus: implications for the pathogenesis of obesity. Milanski M, Degasperi G, Coope A, Morari J, Denis R, Cintra DE, Tsukumo DM, Anhe G, Amaral ME, Takahashi HK, Curi R, Oliveira HC, Carvalheira JB, Bordin S, Saad MJ, Velloso LA. "According to our results, long-chain saturated fatty acids activate predominantly toll-like receptor 4 signaling, which determines not only the induction of local cytokine expression but also promotes endoplasmic reticulum stress. Rats fed on a monounsaturated fat-rich diet do not develop hypothalamic leptin resistance, whereas toll-like receptor 4 loss-of-function mutation and immunopharmacological inhibition of toll-like receptor 4 protects mice from diet-induced obesity"
[58] Biomed Pharmacother. 2010 Oct;64(8):534-40. Glycine regulates inflammatory markers modifying the energetic balance through PPAR and UCP-2. Almanza-Perez JC, Alarcon-Aguilar FJ, Blancas-Flores G, Campos-Sepulveda AE, Roman-Ramos R, Garcia-Macedo R, Cruz M.
[59] Eur J Pharmacol. 2008 Jun 10;587(1-3):317-21. Glycine increases mRNA adiponectin and diminishes pro-inflammatory adipokines expression in 3T3-L1 cells. Garcia-Macedo R, Sanchez-Muñoz F, Almanza-Perez JC, Duran-Reyes G, Alarcon-Aguilar F, Cruz M.
[60] Eur J Pharmacol. 2008 Dec 3;599(1-3):152-8. Glycine regulates the production of pro-inflammatory cytokines in lean and monosodium glutamate-obese mice. Alarcon-Aguilar FJ, Almanza-Perez J, Blancas G, Angeles S, Garcia-Macedo R, Roman R, Cruz M.
[61] Science. 1978 Jul 28;201(4353):358-60. Brain edema: induction in cortical slices by polyunsaturated fatty acids. Chan PH, Fishman RA.
[62] Ann Neurol. 1983 Jun;13(6):625-32. Induction of brain edema following intracerebral injection of arachidonic acid. Chan PH, Fishman RA, Caronna J, Schmidley JW, Prioleau G, Lee J.
[63] Nutr Metab (Lond). 2007 Feb 21;4:4. Long term highly saturated fat diet does not induce NASH in Wistar rats. Romestaing C, Piquet MA, Bedu E, Rouleau V, Dautresme M, Hourmand-Ollivier I, Filippi C, Duchamp C, Sibille B.
[64] Alcohol. 2013 May;47(3):257-64. Ethanol and dietary unsaturated fat (corn oil/linoleic acid enriched) cause intestinal inflammation and impaired intestinal barrier defense in mice chronically fed alcohol. Kirpich IA, Feng W, Wang Y, Liu Y, Beier JI, Arteel GE, Falkner KC, Barve SS, McClain CJ.
[65] Hepatology. 2005 Sep;42(3):568-77. Role of adiponectin in the protective action of dietary saturated fat against alcoholic fatty liver in mice. You M, Considine RV, Leone TC, Kelly DP, Crabb DW.
[66] J Pharmacol Exp Ther. 2001 Nov;299(2):638-44. Dietary saturated fatty acids reverse inflammatory and fibrotic changes in rat liver despite continued ethanol administration. Nanji AA, Jokelainen K, Tipoe GL, Rahemtulla A, Dannenberg AJ.
Ylimääräisiä lähteitä (sekalaista aiheeseen liittyvää tutkimuskirjallisuutta)
[extra] Bol Med Hosp Infant Mex Vol. 67, March-April 2010 Obesity as an inflammatory process. Blancas-Flores G, Almanza-Pérez JC, López-Roa RI, Alarcón-Aguilar FJ, García-Macedo R, Cruz M.
[extra] EMBO Mol Med. 2011 Sep;3(9):559-72. doi: 10.1002/emmm.201100159. Epub 2011 Aug 3. Intestinal mucosal adherence and translocation of commensal bacteria at the early onset of type 2 diabetes: molecular mechanisms and probiotic treatment. Amar J, Chabo C, Waget A, Klopp P, Vachoux C, Bermúdez-Humarán LG, Smirnova N, Bergé M, Sulpice T, Lahtinen S, Ouwehand A, Langella P, Rautonen N, Sansonetti PJ, Burcelin R. "Here, we demonstrate that before the onset of diabetes, after only one week of a high-fat diet (HFD), live commensal intestinal bacteria are present in large numbers in the adipose tissue and the blood where they can induce inflammation. This translocation is prevented in mice lacking the microbial pattern recognition receptors Nod1 or CD14, but overtly increased in Myd88 knockout and ob/ob mouse. [...] The bacterial translocation process from intestine towards tissue can be reversed by six weeks of treatment with the probiotic strain Bifidobacterium animalis subsp. lactis 420, which improves the animals' overall inflammatory and metabolic status."
[extra] PLoS One. 2014 May 14;9(5):e96864. doi: 10.1371/journal.pone.0096864. eCollection 2014. Acute binge drinking increases serum endotoxin and bacterial DNA levels in healthy individuals. Bala S, Marcos M, Gattu A, Catalano D, Szabo G.
[extra] Alcohol Clin Exp Res. 2005 Nov;29(11 Suppl):166S-71S. Activation of the innate immune system and alcoholic liver disease: effects of ethanol per se or enhanced intestinal translocation of bacterial toxins induced by ethanol? Bode C, Bode JC.
[extra] J Hepatol. 1991 Mar;12(2):162-9. Plasma endotoxin concentrations in patients with alcoholic and non-alcoholic liver disease: reevaluation with an improved chromogenic assay. Fukui H, Brauner B, Bode JC, Bode C. "The results indicate that, irrespective of the stage of liver disease, alcohol abuse favours the development of endotoxemia."
[extra] J Hepatol. 2000 May;32(5):742-7. Increased intestinal permeability to macromolecules and endotoxemia in patients with chronic alcohol abuse in different stages of alcohol-induced liver disease. Parlesak A, Schäfer C, Schütz T, Bode JC, Bode C. "Polyethylene glycol Mr 10,000 was detected significantly less frequently in urine from healthy controls (0/30) than in urine of patients with alcoholic liver disease (20/54, [...] Endotoxin concentrations in the plasma of alcoholics were increased more than 5-fold compared to healthy controls [...] The results of this study indicate that alcohol abuse impairs the function of the intestinal barrier, which might enhance the translocation of bacterial toxins, thereby contributing to inflammatory processes in alcoholic liver disease."
[extra] Am J Clin Nutr. 2008 May;87(5):1219-23. Energy intake is associated with endotoxemia in apparently healthy men. Amar J, Burcelin R, Ruidavets JB, Cani PD, Fauvel J, Alessi MC, Chamontin B, Ferriéres J. "In a multivariate analysis, endotoxemia was independently associated with energy intake."
[extra] Diabetes Care. 2011 Feb;34(2):392-7. doi: 10.2337/dc10-1676. Endotoxemia is associated with an increased risk of incident diabetes. Pussinen PJ, Havulinna AS, Lehto M, Sundvall J, Salomaa V.
[extra] Nutr Metab (Lond). 2013 Jan 10;10(1):6. Dietary oil composition differentially modulates intestinal endotoxin transport and postprandial endotoxemia. Mani V, Hollis JH, Gabler NK.
[extra] Exp Physiol. 2014 Jun 27. [Epub ahead of print] Fructose supplementation worsens the deleterious effects of short term high fat feeding on hepatic steatosis and lipid metabolism in adult rats. Crescenzo R, Bianco F, Coppola P, Mazzoli A, Tussellino M, Carotenuto R, Liverini G, Iossa S.
[extra] Alcohol Clin Exp Res. 2012 May;36(5):835-46. doi: 10.1111/j.1530-0277.2011.01673.x. Epub 2011 Dec 7. The type of dietary fat modulates intestinal tight junction integrity, gut permeability, and hepatic toll-like receptor expression in a mouse model of alcoholic liver disease. Kirpich IA, Feng W, Wang Y, Liu Y, Barker DF, Barve SS, McClain CJ. "Compared with control animals, hepatic TLR (TLR 1, 2, 3, 4, 7, 8, 9) mRNA expression was significantly (p < 0.05) increased in USF + EtOH, but not in SF + EtOH group."
[extra] Obesity (Silver Spring). 2011 Nov;19(11):2280-2. doi: 10.1038/oby.2011.251. Epub 2011 Aug 18. Intestinal permeability is associated with visceral adiposity in healthy women. Gummesson A, Carlsson LM, Storlien LH, Bäckhed F, Lundin P, Löfgren L, Stenlöf K, Lam YY, Fagerberg B, Carlsson B.
[extra] Arterioscler Thromb Vasc Biol. 2013 Jun;33(6):1162-70. doi: 10.1161/ATVBAHA.112.300572. Epub 2013 Apr 4. Dietary carnosine prevents early atherosclerotic lesion formation in apolipoprotein E-null mice. Barski OA, Xie Z, Baba SP, Sithu SD, Agarwal A, Cai J, Bhatnagar A, Srivastava S.
[extra] Br J Pharmacol. 2012 Jun;166(4):1344-56. doi: 10.1111/j.1476-5381.2012.01834.x. D-Carnosine octylester attenuates atherosclerosis and renal disease in ApoE null mice fed a Western diet through reduction of carbonyl stress and inflammation. Menini S, Iacobini C, Ricci C, Scipioni A, Blasetti Fantauzzi C, Giaccari A, Salomone E, Canevotti R, Lapolla A, Orioli M, Aldini G, Pugliese G.
[extra] Can J Physiol Pharmacol. 2004 Jul;82(7):506-14. Amino acid metabolism in the Zucker diabetic fatty rat: effects of insulin resistance and of type 2 diabetes. Wijekoon EP, Skinner C, Brosnan ME, Brosnan JT. "At the insulin-resistant stage, the plasma concentrations of the gluconeogenic amino acids aspartate, serine, glutamine, glycine, and histidine were decreased [...] It is evident that insulin resistance alone is capable of bringing about many of the changes in amino acid metabolism observed in type 2 diabetes."
[extra] Br J Nutr. 2012 Jul 14;108(1):57-61. doi: 10.1017/S0007114511005289. Histidine and arginine are associated with inflammation and oxidative stress in obese women. Niu YC, Feng RN, Hou Y, Li K, Kang Z, Wang J, Sun CH, Li Y. "Among the amino acids determined, serum histidine, arginine, threonine, glycine, lysine and serine were found to be significantly lower in obese women as compared to non-obese controls (P < 0·001). The difference was the greatest for histidine (P < 0·001)."
[extra] Gastroenterology. 2009 Feb;136(2):564-74.e2. doi: 10.1053/j.gastro.2008.09.062. Epub 2008 Oct 7. Dietary histidine ameliorates murine colitis by inhibition of proinflammatory cytokine production from macrophages. Andou A, Hisamatsu T, Okamoto S, Chinen H, Kamada N, Kobayashi T, Hashimoto M, Okutsu T, Shimbo K, Takeda T, Matsumoto H, Sato A, Ohtsu H, Suzuki M, Hibi T.
[extra] Am J Physiol Lung Cell Mol Physiol. 2007 May;292(5):L1095-104. Epub 2007 Jan 12. Protective effect of orally administered carnosine on bleomycin-induced lung injury. Cuzzocrea S, Genovese T, Failla M, Vecchio G, Fruciano M, Mazzon E, Di Paola R, Muià C, La Rosa C, Crimi N, Rizzarelli E, Vancheri C.
[extra] Diabetes. 2013 Jul;62(7):2266-77. doi: 10.2337/db12-1701. Epub 2013 Mar 8. Histidine augments the suppression of hepatic glucose production by central insulin action. Kimura K1, Nakamura Y, Inaba Y, Matsumoto M, Kido Y, Asahara S, Matsuda T, Watanabe H, Maeda A, Inagaki F, Mukai C, Takeda K, Akira S, Ota T, Nakabayashi H, Kaneko S, Kasuga M, Inoue H.
[extra] Hum Exp Toxicol. 2010 Aug;29(8):659-65. The effect of carnosine pretreatment on oxidative stress and hepatotoxicity in binge ethanol administered rats. Artun BC, Küskü-Kiraz Z, Güllüoğlu M, Cevikbaş U, Koçak-Toker N, Uysal M. "In conclusion, carnosine prevented the increases in serum transaminase activities and lipid peroxides in liver of ethanol-treated rats, without any change on steatosis in liver."
[extra] Neurosci Lett. 2012 Feb 21;510(1):1-5. Effects of L-carnosine on splenic sympathetic nerve activity and tumor proliferation. Horii Y, Shen J, Fujisaki Y, Yoshida K, Nagai K. "The tumor volumes of the control mice given water gradually and markedly increased and reached a value of 869.8 ± 132.9 mm3 on day 22 after implantation (Fig. 3b). The tumor volumes of the l-carnosine group also increased; however, the increase was less than that of the control group, reaching a value of 407.8 ± 121.7 mm3 (46.9% of the tumor volume of the control group) on day 22."
[extra] Nutr Clin Pract. 2013 Oct;28(5):609-16. doi: 10.1177/0884533613493333. Epub 2013 Jul 8. Effects of L-carnosine and its zinc complex (Polaprezinc) on pressure ulcer healing. Sakae K, Agata T, Kamide R, Yanagisawa H. "After 4 weeks, the rate of pressure ulcer healing, assessed by the mean weekly improvement in PUSH score, was significantly greater in the CAR (1.6 ± 0.2, P = .02) and PLZ groups (1.8 ± 0.2, P = .009) than in the control group (0.8 ± 0.2)."
[extra] J Periodontol. 2002 Jan;73(1):73-8. Systemic release of endotoxins induced by gentle mastication: association with periodontitis severity. Geerts SO, Nys M, De MP, Charpentier J, Albert A, Legrand V, Rompen EH. "For standardizing the mastication session, the patients were asked to gently chew 2 chewing gums 50 times on each side of the jaw under the supervision of a dental practitioner (SG)." "Overall, blood levels of endotoxin after mastication were found to be significantly higher than before mastication (0.89 +/- 3.3 pg/ml versus 3.0 +/- 5.8 pg/ml; P= 0.0002). Likewise, the incidence of positive endotoxemia rose from 6% before mastication to 24% after mastication (P = 0.001)."
[extra] Arterioscler Thromb Vasc Biol. 2004 Nov;24(11):2174-80. Epub 2004 Sep 23. Severe periodontitis enhances macrophage activation via increased serum lipopolysaccharide. Pussinen PJ, Vilkuna-Rautiainen T, Alfthan G, Palosuo T, Jauhiainen M, Sundvall J, Vesanen M, Mattila K, Asikainen S. "LPS concentration correlated positively with serum concentration of oxidized LDL and cytokine production."
[extra] Innate Immun. 2008 Aug;14(4):247-53. doi: 10.1177/1753425908095130. Lipopolysaccharide associates with pro-atherogenic lipoproteins in periodontitis patients. Kallio KA, Buhlin K, Jauhiainen M, Keva R, Tuomainen AM, Klinge B, Gustafsson A, Pussinen PJ. "Endotoxemia increases with severity of periodontitis. In periodontitis, LPS associates preferentially with the pro-atherogenic VLDL-IDL fraction. Periodontal treatment has only minor effects on plasma LPS activity or distribution, which reflects persistence of the disease."
[extra] Intern Med. 2011;50(15):1569-74. Inflammatory cytokines, adiponectin, insulin resistance and metabolic control after periodontal intervention in patients with type 2 diabetes and chronic periodontitis. Sun WL, Chen LL, Zhang SZ, Wu YM, Ren YZ, Qin GM.
[extra] J Clin Periodontol. 2010 Jan;37(1):53-8. Effect of periodontal treatment on metabolic control, systemic inflammation and cytokines in patients with type 2 diabetes. Correa FO1, Gonçalves D, Figueredo CM, Bastos AS, Gustafsson A, Orrico SR.
[extra] FEBS Letters Volume 581, Issue 3, 6 February 2007, Pages 349–354: Augmentation of 11β-hydroxysteroid dehydrogenase type 1 in LPS-activated J774.1 macrophages – Role of 11β-HSD1 in pro-inflammatory properties in macrophages Takako Ishii, Hiroaki Masuzaki, Tomohiro Tanaka, Naoki Arai, Shintaro Yasue, Nozomi Kobayashi, Tsutomu Tomita, Michio Noguchi, Junji Fujikura, Ken Ebihara, Kiminori Hosoda, Kazuwa Nakao
[extra] Endocrinology. 2011 Apr;152(4):1314-26. doi: 10.1210/en.2010-0659. Epub 2011 Jan 25. Low-grade hypothalamic inflammation leads to defective thermogenesis, insulin resistance, and impaired insulin secretion. Arruda AP, Milanski M, Coope A, Torsoni AS, Ropelle E, Carvalho DP, Carvalheira JB, Velloso LA. "In Wistar rats, hypothalamic TNFα blunts the anorexigenic effect of leptin, which is accompanied by reduced leptin signaling and increased expression of suppressor of cytokine signaling 3. In addition, hypothalamic TNFα reduces O(2) consumption and the expression of thermogenic proteins in brown adipose tissue and skeletal muscle. Furthermore, hypothalamic inflammation increases base-line plasma insulin and insulin secretion by isolated pancreatic islets, which is accompanied by an impaired insulin signal transduction in liver and skeletal muscle. Hypothalamic inflammation induced by stearic acid also reduces O(2) consumption and blunts peripheral insulin signal transduction. The use of intracerebroventricular infliximab restores O(2) consumption in obese rats, whereas TNF receptor 1-knockout mice are protected from diet-induced reduced thermogenesis and defective insulin signal transduction. Thus, low-grade inflammation of the hypothalamus is sufficient to induce changes in a number of parameters commonly impaired in obesity and DM2, and TNFα is an important mediator of this process."
[extra] Diabetes. 2012 Jun;61(6):1455-62. doi: 10.2337/db11-0390. Epub 2012 Apr 20. Inhibition of hypothalamic inflammation reverses diet-induced insulin resistance in the liver. Milanski M, Arruda AP, Coope A, Ignacio-Souza LM, Nunez CE, Roman EA, Romanatto T, Pascoal LB, Caricilli AM, Torsoni MA, Prada PO, Saad MJ, Velloso LA. "Obese rodents were treated by intracerebroventricular injections, with immunoneutralizing antibodies against Toll-like receptor (TLR)4 or tumor necrosis factor (TNF)α, and insulin signal transduction, hepatic steatosis, and gluconeogenesis were evaluated. The inhibition of either TLR4 or TNFα reduced hypothalamic inflammation, which was accompanied by the reduction of hypothalamic resistance to leptin and improved insulin signal transduction in the liver. This was accompanied by reduced liver steatosis and reduced hepatic expression of markers of steatosis. Furthermore, the inhibition of hypothalamic inflammation restored defective liver glucose production"
[extra] J Biol Chem. 2009 Dec 25;284(52):36213-22. doi: 10.1074/jbc.M109.030874. Epub 2009 Oct 26. Deletion of tumor necrosis factor-alpha receptor 1 (TNFR1) protects against diet-induced obesity by means of increased thermogenesis. Romanatto T1, Roman EA, Arruda AP, Denis RG, Solon C, Milanski M, Moraes JC, Bonfleur ML, Degasperi GR, Picardi PK, Hirabara S, Boschero AC, Curi R, Velloso LA.
[extra] Diabetologia. 2007 Jun;50(6):1267-76. Epub 2007 Apr 11. C3H/HeJ mice carrying a toll-like receptor 4 mutation are protected against the development of insulin resistance in white adipose tissue in response to a high-fat diet. Poggi M, Bastelica D, Gual P, Iglesias MA, Gremeaux T, Knauf C, Peiretti F, Verdier M, Juhan-Vague I, Tanti JF, Burcelin R, Alessi MC.
[extra] Obesity (Silver Spring). 2008 Jun;16(6):1248-55. doi: 10.1038/oby.2008.210. Epub 2008 Apr 10. Tlr-4 deficiency selectively protects against obesity induced by diets high in saturated fat. Davis JE1, Gabler NK, Walker-Daniels J, Spurlock ME.
[extra] J Nutr Biochem. 2011 Feb;22(2):136-41. doi: 10.1016/j.jnutbio.2009.12.008. Absence of Tlr2 protects against high-fat diet-induced inflammation and results in greater insulin-stimulated glucose transport in cultured adipocytes. Davis JE, Braucher DR, Walker-Daniels J, Spurlock ME.
[extra] Cell Metab. 2009 Oct;10(4):249-59. doi: 10.1016/j.cmet.2009.08.013. MyD88 signaling in the CNS is required for development of fatty acid-induced leptin resistance and diet-induced obesity. Kleinridders A, Schenten D, Könner AC, Belgardt BF, Mauer J, Okamura T, Wunderlich FT, Medzhitov R, Brüning JC. [Wikipedia: "Model organisms have been used in the study of MYD88 function. The gene was originally discovered and cloned by Dan Liebermann and Barbara Hoffman in mice.[3] In that species it is a universal adapter protein as it is used by almost all TLRs (except TLR 3) to activate the transcription factor NF-κB."]
[extra] Circ Res. 2007 Jun 8;100(11):1589-96. Epub 2007 May 3. Toll-like receptor-4 mediates vascular inflammation and insulin resistance in diet-induced obesity. Kim F1, Pham M, Luttrell I, Bannerman DD, Tupper J, Thaler J, Hawn TR, Raines EW, Schwartz MW.
[extra] FASEB J. 2012 Aug;26(8):3493-502. doi: 10.1096/fj.12-208868. Epub 2012 May 16. Timed high-fat diet resets circadian metabolism and prevents obesity. Sherman H, Genzer Y, Cohen R, Chapnik N, Madar Z, Froy O. "Although timed HF-diet-fed mice consumed the same amount of calories as ad libitum low-fat diet-fed mice, they showed 12% reduced body weight, 21% reduced cholesterol levels, and 1.4-fold increased insulin sensitivity. Compared with the HF diet ad libitum, the timed HF diet led to 18% lower body weight, 30% decreased cholesterol levels, 10% reduced TNF-α levels, and 3.7-fold improved insulin sensitivity. Timed HF-diet-fed mice exhibited a better satiated and less stressed phenotype of 25% lower ghrelin and 53% lower corticosterone levels compared with mice fed the timed low-fat diet."
[extra] FASEB J. 2010 Dec;24(12):4948-59. doi: 10.1096/fj.10-164921. Epub 2010 Aug 19. Germ-free C57BL/6J mice are resistant to high-fat-diet-induced insulin resistance and have altered cholesterol metabolism. Rabot S, Membrez M, Bruneau A, Gérard P, Harach T, Moser M, Raymond F, Mansourian R, Chou CJ.
[extra] PLoS One. 2010 Aug 16;5(8):e12191. doi: 10.1371/journal.pone.0012191. High-fat diet: bacteria interactions promote intestinal inflammation which precedes and correlates with obesity and insulin resistance in mouse. Ding S, Chi MM, Scull BP, Rigby R, Schwerbrock NM, Magness S, Jobin C, Lund PK. "Bacteria and HF diet interact to promote proinflammatory changes in the small intestine, which precede weight gain and obesity and show strong and significant associations with progression of obesity and development of insulin resistance. To our knowledge, this is the first evidence that intestinal inflammation is an early consequence of HF diet which may contribute to obesity and associated insulin resistance. Interventions which limit intestinal inflammation induced by HF diet and bacteria may protect against obesity and insulin resistance."
[extra] Diabetes Care. 2011 Aug;34(8):1809-15. doi: 10.2337/dc10-2197. Epub 2011 Jun 2. Bacterial endotoxin activity in human serum is associated with dyslipidemia, insulin resistance, obesity, and chronic inflammation. Lassenius MI, Pietiläinen KH, Kaartinen K, Pussinen PJ, Syrjänen J, Forsblom C, Pörsti I, Rissanen A, Kaprio J, Mustonen J, Groop PH, Lehto M; FinnDiane Study Group.
[extra] Atherosclerosis. 2009 Apr;203(2):494-502. doi: 10.1016/j.atherosclerosis.2008.06.018. Epub 2008 Jul 1. Ethnic and sex differences in circulating endotoxin levels: A novel marker of atherosclerotic and cardiovascular risk in a British multi-ethnic population. Miller MA, McTernan PG, Harte AL, Silva NF, Strazzullo P, Alberti KG, Kumar S, Cappuccio FP. "Endotoxin levels were positively associated with waist, waist-hip ratio, total cholesterol, serum triglycerides and serum insulin levels and negatively associated with serum HDL-cholesterol."
[extra] Dig Dis Sci. 2012 Jul;57(7):1932-41. doi: 10.1007/s10620-012-2112-9. Epub 2012 Mar 17. Nutrition, intestinal permeability, and blood ethanol levels are altered in patients with nonalcoholic fatty liver disease (NAFLD). Volynets V, Küper MA, Strahl S, Maier IB, Spruss A, Wagnerberger S, Königsrainer A, Bischoff SC, Bergheim I. "Despite no differences in the prevalence of bacterial overgrowth and in the orocecal transit time, intestinal permeability, alcohol, and endotoxin levels in plasma were significantly higher in patients with NAFLD than in controls."
[extra] Surgery. 2012 Apr;151(4):587-93. doi: 10.1016/j.surg.2011.09.038. Epub 2011 Nov 16. Reduction in endotoxemia, oxidative and inflammatory stress, and insulin resistance after Roux-en-Y gastric bypass surgery in patients with morbid obesity and type 2 diabetes mellitus. Monte SV, Caruana JA, Ghanim H, Sia CL, Korzeniewski K, Schentag JJ, Dandona P. "LPS, NF-κB DNA binding, TLR-4, TLR-2, and CD14 expression, CRP, MMP-9, and MCP-1 decreased significantly after RYGB. The mechanism underlying resolution of insulin resistance and T2DM after RYGB may be attributable, at least in part, to the reduction of endotoxemia and associated proinflammatory mediators."
[extra] Diabetes Care. 2009 Dec;32(12):2281-7. doi: 10.2337/dc09-0979. Epub 2009 Sep 15. Increase in plasma endotoxin concentrations and the expression of Toll-like receptors and suppressor of cytokine signaling-3 in mononuclear cells after a high-fat, high-carbohydrate meal: implications for insulin resistance. Ghanim H1, Abuaysheh S, Sia CL, Korzeniewski K, Chaudhuri A, Fernandez-Real JM, Dandona P. "HFHC meal intake induced an increase in plasma LPS concentration and the expression of SOCS-3, TLR2, and TLR4 protein, reactive oxygen species generation, and nuclear factor-kappaB binding activity (P < 0.05 for all). These increases were totally absent after the AHA meal rich in fiber and fruit."
[extra] Diabetologia. 2007 Nov;50(11):2374-83. Epub 2007 Sep 6. Selective increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a mechanism associated with endotoxaemia. Cani PD, Neyrinck AM, Fava F, Knauf C, Burcelin RG, Tuohy KM, Gibson GR, Delzenne NM. "Since bifidobacteria have been reported to reduce intestinal endotoxin levels and improve mucosal barrier function, we specifically increased the gut bifidobacterial content of HF-diet-fed mice through the use of a prebiotic (oligofructose [OFS]). [...] Finally, in HF-OFS-treated-mice, Bifidobacterium spp. significantly and positively correlated with improved glucose tolerance, glucose-induced insulin secretion and normalised inflammatory tone (decreased endotoxaemia, plasma and adipose tissue proinflammatory cytokines)."
[extra] Am J Clin Nutr. 2007 Nov;86(5):1286-92. A high-fat meal induces low-grade endotoxemia: evidence of a novel mechanism of postprandial inflammation. Erridge C, Attina T, Spickett CM, Webb DJ. "Baseline endotoxin concentrations were 8.2 pg/mL (interquartile range: 3.4-13.5 pg/mL) but increased significantly (P < 0.05) by approximately 50% after a high-fat meal or after a high-fat meal with cigarettes but not after no meal or cigarettes alone. These results were validated by the observations that a high-fat meal with or without cigarettes, but not no meal or smoking, also significantly (P < 0.05) reduced plasma endotoxin neutralization capacity, which is an indirect measure of endotoxin exposure"
[extra] Atherosclerosis. 2011 May;216(1):1-6. doi: 10.1016/j.atherosclerosis.2011.02.043. Epub 2011 Feb 26. Diet, commensals and the intestine as sources of pathogen-associated molecular patterns in atherosclerosis, type 2 diabetes and non-alcoholic fatty liver disease. Erridge C. "This review summarises the evidence supporting the proposal that 'pathogen-associated molecular patterns' (PAMPs) derived from dietary and commensal sources may contribute to the chronic inflammatory processes that underpin the development of these diseases via stimulation of TLR2 and TLR4."
[extra] Diabetes Care. 2010 May;33(5):991-7. doi: 10.2337/dc09-1630. Epub 2010 Jan 12. Differential effects of cream, glucose, and orange juice on inflammation, endotoxin, and the expression of Toll-like receptor-4 and suppressor of cytokine signaling-3. Deopurkar R, Ghanim H, Friedman J, Abuaysheh S, Sia CL, Mohanty P, Viswanathan P, Chaudhuri A, Dandona P. "Although both glucose and cream induce NF-kappaB binding and an increase in the expression of SOCS3, TNF-alpha, and IL-1beta in MNCs, only cream caused an increase in LPS concentration and TLR-4 expression. Equicaloric amounts of orange juice or water did not induce a change in any of these indexes. These changes are relevant to the pathogenesis of atherosclerosis and insulin resistance."
[extra] Metabolism. 2006 Sep;55(9):1177-85. Glucose ingestion induces an increase in intranuclear nuclear factor kappaB, a fall in cellular inhibitor kappaB, and an increase in tumor necrosis factor alpha messenger RNA by mononuclear cells in healthy human subjects. Aljada A, Friedman J, Ghanim H, Mohanty P, Hofmeyer D, Chaudhuri A, Dandona P.
[extra] Diabetologia. 2007 Feb;50(2):278-85. Epub 2006 Dec 16. Role of inflammatory mediators in the suppression of insulin receptor phosphorylation in circulating mononuclear cells of obese subjects. Ghanim H, Aljada A, Daoud N, Deopurkar R, Chaudhuri A, Dandona P. "We conclude that in obesity the MNC is characterised by reduced p-INSR-beta and increased inflammatory mediators including IKBKB, PRKCB2 and SOCS3. The increase in SOCS3 but not IKBKB or PRKCB2 is related inversely to p-INSR-beta and might mediate the inhibition of p-INSR-beta."
[extra] Gut. 2012 Dec;61(12):1701-7. doi: 10.1136/gutjnl-2011-301689. Epub 2012 Apr 25. Gut-derived lipopolysaccharide augments adipose macrophage accumulation but is not essential for impaired glucose or insulin tolerance in mice. Caesar R1, Reigstad CS, Bäckhed HK, Reinhardt C, Ketonen M, Lundén GÖ, Cani PD, Bäckhed F.
[extra] J Clin Invest. 2003 Dec;112(12):1796-808. Obesity is associated with macrophage accumulation in adipose tissue. Weisberg SP1, McCann D, Desai M, Rosenbaum M, Leibel RL, Ferrante AW Jr. "Adipose tissue macrophage numbers increase in obesity and participate in inflammatory pathways that are activated in adipose tissues of obese individuals."
[extra] FASEB J. 2008 Jul;22(7):2416-26. doi: 10.1096/fj.07-102723. Epub 2008 Mar 7. Gut microbiota modulation with norfloxacin and ampicillin enhances glucose tolerance in mice. Membrez M, Blancher F, Jaquet M, Bibiloni R, Cani PD, Burcelin RG, Corthesy I, Macé K, Chou CJ.
[extra] PLoS One. 2012;7(3):e32967. doi: 10.1371/journal.pone.0032967. Epub 2012 Mar 7. Leptin resistance in vagal afferent neurons inhibits cholecystokinin signaling and satiation in diet induced obese rats. de Lartigue G, Barbier de la Serre C, Espero E, Lee J, Raybould HE.
[extra] Am J Physiol Endocrinol Metab. 2011 Jul;301(1):E187-95. doi: 10.1152/ajpendo.00056.2011. Epub 2011 Apr 26. Diet-induced obesity leads to the development of leptin resistance in vagal afferent neurons. de Lartigue G, Barbier de la Serre C, Espero E, Lee J, Raybould HE.
[extra] MBio. 2014 Jun 10;5(3). pii: e01011-14. doi: 10.1128/mBio.01011-14. Saccharomyces boulardii Administration Changes Gut Microbiota and Reduces Hepatic Steatosis, Low-Grade Inflammation, and Fat Mass in Obese and Type 2 Diabetic db/db Mice. Everard A, Matamoros S, Geurts L, Delzenne NM, Cani PD.
[extra] Diabetes Metab Syndr Obes. 2012;5:175-89. doi: 10.2147/DMSO.S33473. Epub 2012 Jul 6. Comparison with ancestral diets suggests dense acellular carbohydrates promote an inflammatory microbiota, and may be the primary dietary cause of leptin resistance and obesity. Spreadbury I.
[extra] Gastroenterology. 2012 May;142(5):1100-1101.e2. doi: 10.1053/j.gastro.2012.01.034. Epub 2012 Feb 8. A high-fat diet is associated with endotoxemia that originates from the gut. Pendyala S1, Walker JM, Holt PR. "Placing 8 healthy subjects on a Western-style diet for 1 month induced a 71% increase in plasma levels of endotoxin activity (endotoxemia), whereas a prudent-style diet reduced levels by 31%. The Western-style diet might, therefore, contribute to endotoxemia by causing changes in gastrointestinal barrier function or the composition of the microbiota."
[extra] Cell. 2011 Nov 11;147(4):868-80. doi: 10.1016/j.cell.2011.09.051. CD14 controls the LPS-induced endocytosis of Toll-like receptor 4. Zanoni I, Ostuni R, Marek LR, Barresi S, Barbalat R, Barton GM, Granucci F, Kagan JC. "These data establish that upon microbial detection, an upstream PRR (CD14) controls the trafficking and signaling functions of a downstream PRR (TLR4)."
[extra] Ann Surg. 2003 Feb;237(2):246-55. Protective effects of medium-chain triglycerides on the liver and gut in rats administered endotoxin. Kono H, Fujii H, Asakawa M, Yamamoto M, Matsuda M, Maki A, Matsumoto Y. "All rats given corn oil died after LPS administration; however, this mortality was prevented by MCT in a dose-dependent manner. Rats given corn oil showed liver injury after LPS administration. In contrast, MCT prevented this pathologic change nearly completely. MCT blunted CD14 expression on the Kupffer cells and TNF-alpha production by isolated Kupffer cells; however, there were no differences in phagocytic index between the two groups. The length of the intestinal epithelium was increased in the MCT group compared to the corn oil group. Further, after LPS administration, increases in gut permeability and injury were prevented by MCT. Importantly, MCT also prevented hepatic energy charge and gut injuries in this condition."
[extra] Am J Physiol Gastrointest Liver Physiol. 2013 Dec;305(12):G919-32. doi: 10.1152/ajpgi.00226.2013. Epub 2013 Oct 10. Dietary fat sources differentially modulate intestinal barrier and hepatic inflammation in alcohol-induced liver injury in rats. Zhong W, Li Q, Xie G, Sun X, Tan X, Sun X, Jia W, Zhou Z.
[extra] J Gastroenterol. 2013 Aug 17. Toll-like receptor 4 activation in Barrett's esophagus results in a strong increase in COX-2 expression. Verbeek RE, Siersema PD, Ten Kate FJ, Fluiter K, Souza RF, Vleggaar FP, Bus P, van Baal JW.
[extra] Gastroenterology. 2006 Sep;131(3):862-77. Cox-2 is regulated by Toll-like receptor-4 (TLR4) signaling: Role in proliferation and apoptosis in the intestine. Fukata M, Chen A, Klepper A, Krishnareddy S, Vamadevan AS, Thomas LS, Xu R, Inoue H, Arditi M, Dannenberg AJ, Abreu MT.
[extra] J Leukoc Biol. 2009 Oct;86(4):971-80. doi: 10.1189/jlb.0708396. Epub 2009 Jun 29. TLR4-mediated Cox-2 expression increases intestinal ischemia/reperfusion-induced damage. Moses T, Wagner L, Fleming SD.
[extra] BMC Gastroenterol. 2010 Jul 16;10:82. doi: 10.1186/1471-230X-10-82. The role of prostaglandin E2 (PGE 2) in toll-like receptor 4 (TLR4)-mediated colitis-associated neoplasia. Hernandez Y, Sotolongo J, Breglio K, Conduah D, Chen A, Xu R, Hsu D, Ungaro R, Hayes LA, Pastorini C, Abreu MT, Fukata M.
[extra] Inflamm Bowel Dis. 2011 Jul;17(7):1464-73. Constitutive activation of epithelial TLR4 augments inflammatory responses to mucosal injury and drives colitis-associated tumorigenesis. Fukata M1, Shang L, Santaolalla R, Sotolongo J, Pastorini C, España C, Ungaro R, Harpaz N, Cooper HS, Elson G, Kosco-Vilbois M, Zaias J, Perez MT, Mayer L, Vamadevan AS, Lira SA, Abreu MT.
[extra] J Immunol. 2009 Feb 15;182(4):2476-84. doi: 10.4049/jimmunol.0802059. Lactate boosts TLR4 signaling and NF-kappaB pathway-mediated gene transcription in macrophages via monocarboxylate transporters and MD-2 up-regulation. Samuvel DJ1, Sundararaj KP, Nareika A, Lopes-Virella MF, Huang Y.
[extra] J Neurosci. 2010 Jun 16;30(24):8285-95. doi: 10.1523/JNEUROSCI.0976-10.2010. Pivotal role of TLR4 receptors in alcohol-induced neuroinflammation and brain damage. Alfonso-Loeches S, Pascual-Lucas M, Blanco AM, Sanchez-Vera I, Guerri C.
[extra] Front Cell Neurosci. 2014 May 27;8:146. doi: 10.3389/fncel.2014.00146. eCollection 2014. TLR4-mediated brain inflammation halts neurogenesis: impact of hormonal replacement therapy. Mouihate A. "These data strongly suggest that combined 17β-estradiol and progesterone, and not 17β-estradiol alone, rescues neurogenesis from the deleterious effect of brain inflammation likely via the inhibition of the signaling pathways leading to the activation of proinflammatory genes."
[extra] Wiad Lek. 2013;66(1):3-9. [The role of TLR4 receptor in development of inflammation and carcinogenesis in ulcerative colitis and pharmacotherapy of this disorder]. [Article in Polish] Szumilas D1, Krysiak R, Okopień B. "The increased expression of TLR4 and the development of the uncontrolled inflammatory response in UC (increased production of COX-2, PGE2, TNFalpha and CCL2) impairs regeneration of the mucosa, resulting in its damage, and may later lead to the development of colon cancer."
[extra] AAPS J. 2014 Mar;16(2):246-57. doi: 10.1208/s12248-013-9558-3. Epub 2014 Jan 15. Antitumor activity of gemcitabine can be potentiated in pancreatic cancer through modulation of TLR4/NF-κB signaling by 6-shogaol. Zhou L, Qi L, Jiang L, Zhou P, Ma J, Xu X, Li P. "Overall, our results suggest that 6-shogaol can inhibit the growth of human pancreatic tumors and sensitize them to gemcitabine by suppressing of TLR4/NF-κB-mediated inflammatory pathways linked to tumorigenesis."
[extra] Inflammation. 2014 Apr;37(2):467-77. doi: 10.1007/s10753-013-9760-6. Saturated fatty acids up-regulate COX-2 expression in prostate epithelial cells via toll-like receptor 4/NF-κB signaling. Liu J, Hu S, Cui Y, Sun MK, Xie F, Zhang Q, Jin J.
[extra] Biochem Biophys Res Commun. 2013 Aug 23;438(2):249-56. doi: 10.1016/j.bbrc.2013.07.006. Epub 2013 Jul 11. Differential COX-2 induction by viral and bacterial PAMPs: Consequences for cytokine and interferon responses and implications for anti-viral COX-2 directed therapies. Kirkby NS, Zaiss AK, Wright WR, Jiao J, Chan MV, Warner TD, Herschman HR, Mitchell JA. "LPS induced Cox2 expression in all tissues examined. In contrast, poly(I:C) elicited a milder response, limited to a subset of tissues."
[extra] J Dent Res. 2009 Jun;88(6):519-23. doi: 10.1177/0022034509338353. Is obesity an oral bacterial disease? Goodson JM, Groppo D, Halem S, Carpino E. "Classification tree analysis of salivary microbiological composition revealed that 98.4% of the overweight women could be identified by the presence of a single bacterial species (Selenomonas noxia) at levels greater than 1.05% of the total salivary bacteria."
[extra] Aging Cell. 2013 Dec;12(6):1041-9. doi: 10.1111/acel.12133. Epub 2013 Aug 20. The phytochemical, EGCG, extends lifespan by reducing liver and kidney function damage and improving age-associated inflammation and oxidative stress in healthy rats. Niu Y, Na L, Feng R, Gong L, Zhao Y, Li Q, Li Y, Sun C.
[extra] Biochem Biophys Res Commun. 2012 Oct 5;426(4):480-5. doi: 10.1016/j.bbrc.2012.08.096. Epub 2012 Aug 29. Green tea polyphenol epigallocatechin-3-gallate inhibits TLR4 signaling through the 67-kDa laminin receptor on lipopolysaccharide-stimulated dendritic cells. Byun EB, Choi HG, Sung NY, Byun EH.
[extra] J Immunol. 2010 Jul 1;185(1):33-45. doi: 10.4049/jimmunol.0903742. Epub 2010 May 28. TLR4 signaling inhibitory pathway induced by green tea polyphenol epigallocatechin-3-gallate through 67-kDa laminin receptor. Hong Byun E1, Fujimura Y, Yamada K, Tachibana H.
[extra] Mediators Inflamm. 2009;2009:704706. doi: 10.1155/2009/704706. Epub 2010 Feb 16. Oxymatrine downregulates TLR4, TLR2, MyD88, and NF-kappaB and protects rat brains against focal ischemia. Fan H, Li L, Zhang X, Liu Y, Yang C, Yang Y, Yin J. "The excessive inflammatory immune reaction often resides in region of necrosis and ischemic tissue after cerebral infarction and leads to inflammatory injury. The relationship between toll-like receptor-4 (TLR4), toll-like receptor-2 (TLR2), and nonbiological inflammatory injury has been proved [1–3]. It is believed that injured tissue and necrotic cells release endogenous activators (adjuvants). These activators can combine with TLR4 in the cell membrane. [...] An excessive inflammatory reaction can also damage target cells and tissue [4, 5]. Reducing the inflammatory injury is therefore regarded to be one of major ways to treat acute cerebral infarction."
[extra] Stroke. 2013 Jan;44(1):205-12. Safety and efficacy evaluation of carnosine, an endogenous neuroprotective agent for ischemic stroke. Bae ON, Serfozo K, Baek SH, Lee KY, Dorrance A, Rumbeiha W, Fitzgerald SD, Farooq MU, Naravelta B, Bhatt A, Majid A. "In both permanent and transient ischemic models, carnosine treatment exhibited significant cerebroprotection against histological and functional damage, with wide therapeutic and clinically relevant time windows. Carnosine was well tolerated and exhibited no toxicity."
[extra] Rejuvenation Res. 2008 Feb;11(1):201-14. doi: 10.1089/rej.2007.0608. Dietary supplementation exerts neuroprotective effects in ischemic stroke model. Yasuhara T, Hara K, Maki M, Masuda T, Sanberg CD, Sanberg PR, Bickford PC, Borlongan CV.
[extra] Cardiovasc Res. 2004 Feb 15;61(3):538-47. Modulating Toll-like receptor mediated signaling by (1-->3)-beta-D-glucan rapidly induces cardioprotection. Li C, Ha T, Kelley J, Gao X, Qiu Y, Kao RL, Browder W, Williams DL. "GP treatment reduced infarct size by 47% in rat hearts subjected to reperfusion for 4 h"
[extra] Gut. 2007 Feb;56(2):168-75. Epub 2006 Jun 15. Zinc carnosine, a health food supplement that stabilises small bowel integrity and stimulates gut repair processes. Mahmood A, FitzGerald AJ, Marchbank T, Ntatsaki E, Murray D, Ghosh S, Playford RJ.
[extra] J Clin Biochem Nutr. 2010 May;46(3):234-43. doi: 10.3164/jcbn.09-125. Epub 2010 Apr 10. Polaprezinc Protects Mice against Endotoxin Shock. Ohata S, Moriyama C, Yamashita A, Nishida T, Kusumoto C, Mochida S, Minami Y, Nakada J, Shomori K, Inagaki Y, Ohta Y, Matsura T. "PZ pretreatment inhibited the decrease in the survival rate of mice after LPS injection. PZ inhibited the increases in plasma NO as well as TNF-alpha after LPS. Compatibly, PZ suppressed LPS-induced inducible NO synthase mRNA transcription in the mouse lungs. PZ also improved LPS-induced lung injury."
[extra] ISME J. 2013 Apr;7(4):880-4. doi: 10.1038/ismej.2012.153. Epub 2012 Dec 13. An opportunistic pathogen isolated from the gut of an obese human causes obesity in germfree mice. Fei N, Zhao L.
[extra] J Am Coll Cardiol. 1999 Dec;34(7):1975-81. Association of endotoxemia with carotid atherosclerosis and cardiovascular disease: prospective results from the Bruneck Study. Wiedermann CJ1, Kiechl S, Dunzendorfer S, Schratzberger P, Egger G, Oberhollenzer F, Willeit J. "Median endotoxin concentration amounted to 14.3 pg/ml (range, 6.0 to 209.2 pg/ml). Subjects with levels beyond 50 pg/ml (90th percentile) faced a threefold risk of incident atherosclerosis (odds ratio [95% confidence interval] 2.9 [1.4-6.3]; p < 0.01). The risk associated with high endotoxin was most pronounced in subjects with chronic infections and in current and ex-smokers."
[extra] Arterioscler Thromb Vasc Biol. 2007 Jun;27(6):1433-9. Epub 2007 Mar 15. Endotoxemia, immune response to periodontal pathogens, and systemic inflammation associate with incident cardiovascular disease events. Pussinen PJ, Tuomisto K, Jousilahti P, Havulinna AS, Sundvall J, Salomaa V. "The corresponding HRs for high endotoxin concentration were 1.82 (1.22 to 2.73, alone), 3.92 (1.99 to 7.74, with CRP), 3.54 (1.78 to 7.03, with IL-6), and 2.26 (1.13 to 4.52, with tumor necrosis factor (TNF)-alpha) after adjusting for age and gender. These associations were abolished after adjusting for serum lipids."
[extra] Arterioscler Thromb Vasc Biol. 2004 Dec;24(12):2227-36. Epub 2004 Oct 7. Potential role of endotoxin as a proinflammatory mediator of atherosclerosis. Stoll LL, Denning GM, Weintraub NL. "In this article, we outline the main elements of the endotoxin signaling receptor complex that initiates proinflammatory signaling (lipopolysaccharide binding protein [LBP], CD14, TLR-4, and MD-2) and discuss how changes in expression of these molecules may affect proatherogenic responses in the vessel wall. We also describe some of the proinflammatory effects of endotoxin that may be relevant to atherosclerosis, and discuss how serum lipoproteins, especially high-density lipoprotein, may modulate endotoxin-induced inflammatory responses. Further, we discuss recent findings suggesting that the lipid-lowering statins may have an additional protective role in blocking at least some of these proinflammatory signaling pathways."
[extra] Metab Brain Dis. 2014 Mar;29(1):19-36. doi: 10.1007/s11011-013-9435-x. Epub 2013 Sep 10. Mitochondrial dysfunctions in myalgic encephalomyelitis/chronic fatigue syndrome explained by activated immuno-inflammatory, oxidative and nitrosative stress pathways. Morris G, Maes M. "Increased levels of pro-inflammatory cytokines, such as interleukin-1 and tumor necrosis factor-α, and elastase, and increased O&NS may inhibit mitochondrial respiration, decrease the activities of the electron transport chain and mitochondrial membrane potential, increase mitochondrial membrane permeability, interfere with ATP production and cause mitochondrial shutdown. The activated O&NS pathways may additionally lead to damage of mitochondrial DNA and membranes thus decreasing membrane fluidity. Lowered levels of antioxidants, zinc and coenzyme Q10, and ω3 polyunsaturated fatty acids in ME/cfs may further aggravate the activated immuno-inflammatory and O&NS pathways."hi
[extra] Neurochem Res. 2005 Oct;30(10):1283-8. Carnosine protects the brain of rats and Mongolian gerbils against ischemic injury: after-stroke-effect. Dobrota D, Fedorova T, Stvolinsky S, Babusikova E, Likavcanova K, Drgova A, Strapkova A, Boldyrev A. "In this study, the therapeutic effect of carnosine was estimated on animals when this drug was administered intraperitoneally (100 mg/kg body weight) after ischemic episode induced by experimental global brain ischemia. Treatment of the animals with carnosine after ischemic episode under long-term (7-14 days) reperfusion demonstrated its pronounced protective effect on neurological symptoms and animal mortality. Carnosine also prevented higher lipid peroxidation of brain membrane structures and increased a resistance of neuronal membranes to the in vitro induced oxidation. Measurements of malonyl dialdehyde (MDA) in brain homogenates showed its increase in the after brain stroke animals and decreased MDA level in the after brain stroke animals treated with carnosine."
[extra] Arch Gerontol Geriatr. 2014 May 2. Anserine and carnosine supplementation in the elderly: Effects on cognitive functioning and physical capacity. Szcześniak D, Budzeń S, Kopeć W, Rymaszewska J. [13 weeks, 0.33g/d carnosine, 0.66g/d anserine -> BMI decreased by 0.51]
[extra] Journal of Human Kinetics, Volume 40, Issue 1 (Mar 2014). Effect of Choline Supplementation on Rapid Weight Loss and Biochemical Variables Among Female Taekwondo and Judo Athlete. Gehan Elsawy, Osama Abdelrahman, Amr Hamza. [significantly increased fat loss and decreased MDA with 4g/d choline]
[extra] Diabetes. 2006 Sep;55(9):2554-61. Exercise improves insulin and leptin sensitivity in hypothalamus of Wistar rats. Flores MB, Fernandes MF, Ropelle ER, Faria MC, Ueno M, Velloso LA, Saad MJ, Carvalheira JB.
[extra] PLoS Biol. 2010 Aug 24;8(8). pii: e1000465. doi: 10.1371/journal.pbio.1000465. IL-6 and IL-10 anti-inflammatory activity links exercise to hypothalamic insulin and leptin sensitivity through IKKbeta and ER stress inhibition. Ropelle ER, Flores MB, Cintra DE, Rocha GZ, Pauli JR, Morari J, de Souza CT, Moraes JC, Prada PO, Guadagnini D, Marin RM, Oliveira AG, Augusto TM, Carvalho HF, Velloso LA, Saad MJ, Carvalheira JB.
[extra] Mol Cell Biol. 2014 Jul 28. pii: MCB.00157-14. [Epub ahead of print] 12-Lipoxygenase Promotes Obesity-Induced Oxidative Stress in Pancreatic Islets. Tersey SA, Maier B, Nishiki Y, Maganti AV, Nadler JL, Mirmira RG. "High fat diets lead to obesity, inflammation, and dysglycemia. 12-lipoxygenase (12-LO) is activated by high fat diets and catalyzes the oxygenation of cellular arachidonic acid to form proinflammatory intermediates. [...] Whereas knockout mice and control littermates displayed identical weight gain, body fat distribution, and macrophage infiltration into fat, knockout mice exhibited greater adaptive islet hyperplasia, improved insulin secretion, and complete protection from dysglycemia."
[extra] Mol Cell Biochem. 2009 Feb;322(1-2):127-35. doi: 10.1007/s11010-008-9949-4. Epub 2008 Nov 11. Curcumin attenuates inflammation through inhibition of TLR-4 receptor in experimental colitis. Lubbad A, Oriowo MA, Khan I. "TNBS-induced increase in the level of MPO activity and MDA concentrations was reversed by curcumin treatment, whereas the same dose of curcumin did not affect their levels in the non-colitis animals. Increases in the levels of TLR-4, MyD88, and NFkB proteins in inflamed tissue were also suppressed significantly by curcumin treatment."
[extra] Cell. 2014 Aug 14;158(4):705-21. doi: 10.1016/j.cell.2014.05.052. Altering the Intestinal Microbiota during a Critical Developmental Window Has Lasting Metabolic Consequences. Cox LM, Yamanishi S, Sohn J, Alekseyenko AV, Leung JM, Cho I, Kim SG, Li H, Gao Z, Mahana D, Zárate Rodriguez JG, Rogers AB, Robine N, Loke P, Blaser MJ. "LDP that is limited to early life transiently perturbs the microbiota, which is sufficient to induce sustained effects on body composition, indicating that microbiota interactions in infancy may be critical determinants of long-term host metabolic effects. In addition, LDP enhances the effect of high-fat diet induced obesity. The growth promotion phenotype is transferrable to germ-free hosts by LDP-selected microbiota, showing that the altered microbiota, not antibiotics per se, play a causal role."
[extra] Diabetes Care. 2008 Sep;31(9):1827-31. doi: 10.2337/dc08-0561. Epub 2008 Jun 12. Acute modulation of toll-like receptors by insulin. Ghanim H, Mohanty P, Deopurkar R, Sia CL, Korzeniewski K, Abuaysheh S, Chaudhuri A, Dandona P.
[extra] Diabetes Care. 2010 Nov;33(11):2416-23. doi: 10.2337/dc10-0929. Epub 2010 Aug 10. Insulin suppresses endotoxin-induced oxidative, nitrosative, and inflammatory stress in humans. Dandona P, Ghanim H, Bandyopadhyay A, Korzeniewski K, Ling Sia C, Dhindsa S, Chaudhuri A. "The coinfusion of insulin led to a total elimination of the increase in NOM, free fatty acids, and TBARS and a significant reduction in ROS generation by PMNLs and plasma MIF, visfatin, and myoglobin concentrations. Insulin did not affect TNF-α, MCP-1, IL-6, LBP, resistin, and HMG-B1 increases induced by the LPS."
[extra] Oncogene. 2014 Jan 2;33(1):74-84. doi: 10.1038/onc.2012.540. Epub 2012 Nov 26. Enhancement of leptin receptor signaling by SOCS3 deficiency induces development of gastric tumors in mice. Inagaki-Ohara K, Mayuzumi H, Kato S, Minokoshi Y, Otsubo T, Kawamura YI, Dohi T, Matsuzaki G, Yoshimura A. "These data suggest that SOCS3 is an antigastric tumor gene that suppresses leptin overexpression and ObRb/STAT3 hyperactivation, supporting the hypothesis that the leptin/ObRb/STAT3 axis accelerates tumorigenesis and that it may represent a new therapeutic target for the treatment of gastric cancer."
[extra] Nat Med. 2004 Jul;10(7):734-8. Epub 2004 Jun 27. Enhanced leptin sensitivity and attenuation of diet-induced obesity in mice with haploinsufficiency of Socs3. Howard JK, Cave BJ, Oksanen LJ, Tzameli I, Bjørbaek C, Flier JS.
[extra] Nat Med. 2004 Jul;10(7):739-43. Epub 2004 Jun 20. Socs3 deficiency in the brain elevates leptin sensitivity and confers resistance to diet-induced obesity. Mori H, Hanada R, Hanada T, Aki D, Mashima R, Nishinakamura H, Torisu T, Chien KR, Yasukawa H, Yoshimura A. "These data indicate that Socs3 is a key regulator of diet-induced leptin as well as insulin resistance. Our study demonstrates the negative regulatory role of Socs3 in leptin signaling in vivo, and thus suppression of Socs3 in the brain is a potential therapy for leptin-resistance in obesity."
[extra] Dev Cell. 2002 Apr;2(4):489-95. PTP1B regulates leptin signal transduction in vivo. Zabolotny JM, Bence-Hanulec KK, Stricker-Krongrad A, Haj F, Wang Y, Minokoshi Y, Kim YB, Elmquist JK, Tartaglia LA, Kahn BB, Neel BG.
[extra] Am J Physiol Endocrinol Metab. 2013 Sep 1;305(5):E649-59. Acute exercise suppresses hypothalamic PTP1B protein level and improves insulin and leptin signaling in obese rats. Chiarreotto-Ropelle EC, Pauli LS, Katashima CK, Pimentel GD, Picardi PK, Silva VR, de Souza CT, Prada PO, Cintra DE, Carvalheira JB, Ropelle ER, Pauli JR. "Interestingly, the anti-inflammatory action and the reduction of PTP1B activity mediated by exercise occurred in an interleukin-6 (IL-6)-dependent manner because exercise failed to reduce inflammation and PTP1B protein level after the disruption of hypothalamic-specific IL-6 action in obese rats. Conversely, intracerebroventricular administration of recombinant IL-6 reproduced the effects of exercise, improving hypothalamic insulin and leptin action by reducing the inflammatory signaling and PTP1B activity in obese rats at rest. Taken together, our study reports that physical exercise restores insulin and leptin signaling, at least in part, by reducing hypothalamic PTP1B protein level through the central anti-inflammatory response."
[extra] Nat Med. 2006 Aug;12(8):917-24. Epub 2006 Jul 16. Neuronal PTP1B regulates body weight, adiposity and leptin action. Bence KK, Delibegovic M, Xue B, Gorgun CZ, Hotamisligil GS, Neel BG, Kahn BB. "Thus, PTP1B regulates body mass and adiposity primarily through actions in the brain. Furthermore, neuronal PTP1B regulates adipocyte leptin production and probably is essential for the development of leptin resistance."
[extra] Cell. 2008 Oct 3;135(1):61-73. doi: 10.1016/j.cell.2008.07.043. Hypothalamic IKKbeta/NF-kappaB and ER stress link overnutrition to energy imbalance and obesity. Zhang X, Zhang G, Zhang H, Karin M, Bai H, Cai D. "Overnutrition atypically activates hypothalamic IKKbeta/NF-kappaB at least in part through elevated endoplasmic reticulum stress in the hypothalamus. While forced activation of hypothalamic IKKbeta/NF-kappaB interrupts central insulin/leptin signaling and actions, site- or cell-specific suppression of IKKbeta either broadly across the brain or locally within the mediobasal hypothalamus, or specifically in hypothalamic AGRP neurons significantly protects against obesity and glucose intolerance. The molecular mechanisms involved include regulation by IKKbeta/NF-kappaB of SOCS3, a core inhibitor of insulin and leptin signaling."
[extra] J Clin Invest. 2009 Sep;119(9):2577-89. doi: 10.1172/JCI36714. Epub 2009 Aug 10. Palmitic acid mediates hypothalamic insulin resistance by altering PKC-theta subcellular localization in rodents. Benoit SC, Kemp CJ, Elias CF, Abplanalp W, Herman JP, Migrenne S, Lefevre AL, Cruciani-Guglielmacci C, Magnan C, Yu F, Niswender K, Irani BG, Holland WL, Clegg DJ. "CNS exposure to palmitic acid via direct infusion or by oral gavage increased the localization of PKC-theta to cell membranes in the hypothalamus, which was associated with impaired hypothalamic insulin and leptin signaling. This finding was specific for palmitic acid, as the monounsaturated fatty acid, oleic acid, neither increased membrane localization of PKC-theta nor induced insulin resistance. Finally, arcuate-specific knockdown of PKC-theta attenuated diet-induced obesity and improved insulin signaling. These results suggest that many of the deleterious effects of high-fat diets, specifically those enriched with palmitic acid, are CNS mediated via PKC-theta activation, resulting in reduced insulin activity."
[extra] Diabetes. 1999 Jun;48(6):1270-4. Free fatty acid-induced insulin resistance is associated with activation of protein kinase C theta and alterations in the insulin signaling cascade. Griffin ME, Marcucci MJ, Cline GW, Bell K, Barucci N, Lee D, Goodyear LJ, Kraegen EW, White MF, Shulman GI. "We conclude that acute elevations of plasma FFA levels for 5 h induce skeletal muscle insulin resistance in vivo via a reduction in insulin-stimulated muscle glycogen synthesis and glucose oxidation that can be attributed to reduced glucose transport activity. These changes are associated with abnormalities in the insulin signaling cascade and may be mediated by FFA activation of PKC theta."
[extra] Mol Immunol. 2009 Dec;47(2-3):420-5. doi: 10.1016/j.molimm.2009.08.023. Epub 2009 Sep 20. Molecular mechanism underlying the suppression of lipid oxidation during endotoxemia. Maitra U, Chang S, Singh N, Li L. "In this report, we have demonstrated that LPS suppresses free fatty acid (FFA) oxidation, and consequently contributes to elevated plasma levels of FFA and triglyceride (TG). Furthermore, this process depends upon the interleukin-1 receptor associated kinase 1 (IRAK-1), one of the key TLR4 intracellular signaling kinases. IRAK-1(-/-) mice fail to exhibit the dramatic rise in plasma FFA and TG levels compared to wild-type (WT) mice following lethal LPS injection. Mechanistically, we demonstrated that LPS suppresses FFA oxidation through decreasing the expression levels of key FFA oxidative genes including CPT-1 and MCAD in both liver and kidney tissues of WT but not IRAK-1(-/-) mice."
[extra] Mol Immunol. 2012 Apr;50(4):244-52. Reduced oxidative tissue damage during endotoxemia in IRAK-1 deficient mice. Singh N, Li L. "In contrast, LPS challenge failed to reduce the activities of GPx and SOD in IRAK-1 deficient tissues."
[extra] Chang, Samantha, and Liwu Li. "Metabolic endotoxemia: a novel concept in chronic disease pathology." J Med Sci 3 (2011): 191-209.
[extra] Hepatology. 2003 Mar;37(3):551-7. Oral bile acids reduce bacterial overgrowth, bacterial translocation, and endotoxemia in cirrhotic rats. Lorenzo-Zúñiga V, Bartolí R, Planas R, Hofmann AF, Viñado B, Hagey LR, Hernández JM, Mañé J, Alvarez MA, Ausina V, Gassull MA.
[extra] Proc Natl Acad Sci U S A. 2004 Nov 2;101(44):15718-23. The gut microbiota as an environmental factor that regulates fat storage. Bäckhed F, Ding H, Wang T, Hooper LV, Koh GY, Nagy A, Semenkovich CF, Gordon JI. "We found that conventionalization of adult germ-free (GF) C57BL/6 mice with a normal microbiota harvested from the distal intestine (cecum) of conventionally raised animals produces a 60% increase in body fat content and insulin resistance within 14 days despite reduced food intake."
[extra] ISME J. 2013 Apr;7(4):880-4. doi: 10.1038/ismej.2012.153. Epub 2012 Dec 13. An opportunistic pathogen isolated from the gut of an obese human causes obesity in germfree mice. Fei N, Zhao L."Here we show that one endotoxin-producing bacterium isolated from a morbidly obese human's gut induced obesity and insulin resistance in germfree mice. The endotoxin-producing Enterobacter decreased in relative abundance from 35% of the volunteer's gut bacteria to non-detectable, during which time the volunteer lost 51.4 kg of 174.8 kg initial weight and recovered from hyperglycemia and hypertension after 23 weeks on a diet of whole grains, traditional Chinese medicinal foods and prebiotics. [...] Mono-association of germfree C57BL/6J mice with strain Enterobacter cloacae B29 isolated from the volunteer's gut induced fully developed obesity and insulin resistance on a high-fat diet but not on normal chow diet, whereas the germfree control mice on a high-fat diet did not exhibit the same disease phenotypes."
[extra] Gastroenterology. 2012 Oct;143(4):913-6.e7. Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Vrieze A, Van Nood E, Holleman F, Salojärvi J, Kootte RS, Bartelsman JF, Dallinga-Thie GM, Ackermans MT, Serlie MJ, Oozeer R, Derrien M, Druesne A, Van Hylckama Vlieg JE, Bloks VW, Groen AK, Heilig HG, Zoetendal EG, Stroes ES, de Vos WM, Hoekstra JB, Nieuwdorp M. "Subjects were assigned randomly to groups that were given small intestinal infusions of allogenic or autologous microbiota. Six weeks after infusion of microbiota from lean donors, insulin sensitivity of recipients increased (median rate of glucose disappearance changed from 26.2 to 45.3 μmol/kg/min; P < .05) along with levels of butyrate-producing intestinal microbiota."
Hyvä Valtsu! :)
VastaaPoistaMitahan siniset - 455-470 nm - valon aallonpituudet voi olla apuna tossa tulehduksessa? Ittella on kokemuksia vain lokaalin inflammaation hoidossa, esim bursiitti tai tendovaginiitti etc. Todella tehokas hoito akuutissa inflammaatiossa.
VastaaPoistaItse toistaiseksi uskon, että sininen valo toimii lähinnä kudoksen pinnalla (esim aknen hoidossa), sillä eihän se optisten ominaisuuksiensa vuoksi läpäise kudosta juuri yhtään.
Poista